
Покрытосеменные растения — основа биосферы
Покрытосеменные (или цветковые) — высшие представители флоры. Это самые распространённые растения на планете. Например, к ним относятся пшеница, дуб, яблоня, пальма, крапива и многие другие. Виды из этого отдела разнообразны по форме, размерам и условиям обитания. Но всех их объединяют общие признаки строения, развития и размножения. О них и пойдёт речь в этом сообщении.
Строение и особенности, отличия от голосеменных
Основные органы, которые есть у любого цветкового растения:
- корень, с помощью которого оно получает питательные вещества из почвы;
- стебель и расположенные на нём боковые побеги;
- листья, отвечающие за фотосинтез;
- цветок — укороченный побег, который со временем меняет свой внешний вид и превращается в плод.
Схема строения цветка.
Несколько важных отличий от голосеменных:
- Орган семенного размножения — цветок, в котором происходит оплодотворение.

- Семена цветковых растений покрыты тканями плода. У голосеменных семя открыто лежит на чешуе шишки. За это они и получили свои основные названия: «покрытосеменные» и «голосеменные»
- Листья могут быть разной формы. Листва голосеменных — это только хвоя.
- Разнообразные жизненные формы — деревья, кустарники и травы. Голосеменные представлены только древесными формами.
Пшеница — представитель покрытосеменных растений. Из пшеничной муки пекут хлеб, делают макароны, пирожные и торты.
Фото: flickr.com/SleepyClaus
Виды и среда обитания
В отделе покрытосеменных насчитывается более 240 000 видов. В биологии всё их многообразие принято делить на два класса: однодольные (рожь, пшеница) и двудольные (бобы, картофель). Классы в свою очередь подразделяют на семейства.
Горох представляет собой слегка вьющуюся траву. Его выращивают почти в каждом огороде, и им любят лакомиться дети.
Фото: flickr.com/LisaMoffatt
За долгое время эволюции цветковые растения приспособились к различной среде обитания — от тропических лесов до пустынь и тундры. Сейчас они широко распространены во всех климатических поясах Земли.
Происхождение
Покрытосеменные и голосеменные произошли от одного общего предка — древних водорослей. Затем в процессе эволюции их пути разделились. У покрытосеменных появился новый орган размножения — цветок, который привлекает насекомых сильным запахом и яркой окраской. Они переносят пыльцу от одного цветка к другому. Так обеспечивается более надёжное опыление растения не только ветром, как у голосеменных, но и насекомыми.
Поэтому покрытосеменные быстро распространились по всей планете и заняли господствующее положение.
Размножение и продолжительность жизни
Цветковые в основном размножаются семенами.
Сначала внутри цветка семя созревает и накапливает питательные вещества. После попадания пыльцы в цветок происходит оплодотворение. Семя постепенно превращается в плод. Внутри него образуются новые семена, которые пока надёжно защищены тканями плода от преждевременного распространения. Когда придёт время, семя упадёт на землю и прорастёт в новое растение.
После попадания пыльцы в цветок происходит оплодотворение. Семя постепенно превращается в плод. Внутри него образуются новые семена, которые пока надёжно защищены тканями плода от преждевременного распространения. Когда придёт время, семя упадёт на землю и прорастёт в новое растение.
Размножение цветковых растений.
Также цветковые могут размножаться побегами и корнями.
По продолжительности жизненного цикла они могут быть:
- однолетними,
- двулетними,
- многолетними.
Независимо от срока своей жизни, каждое покрытосеменное растение обязательно цветёт хотя бы один раз.
Цветущая яблоня радует глаз и источает чудесный аромат, а спелые яблоки полезны для здоровья.
Фото: flickr.com/narujen
Роль в природе и жизни человека
Окружающую нас среду невозможно представить без цветковых растений. В природе они образуют целые ландшафты. В России это леса, степи, лесостепи, тундра.
Они участвуют в круговороте веществ в природе — поглощают из атмосферы углекислый газ и вырабатывают кислород.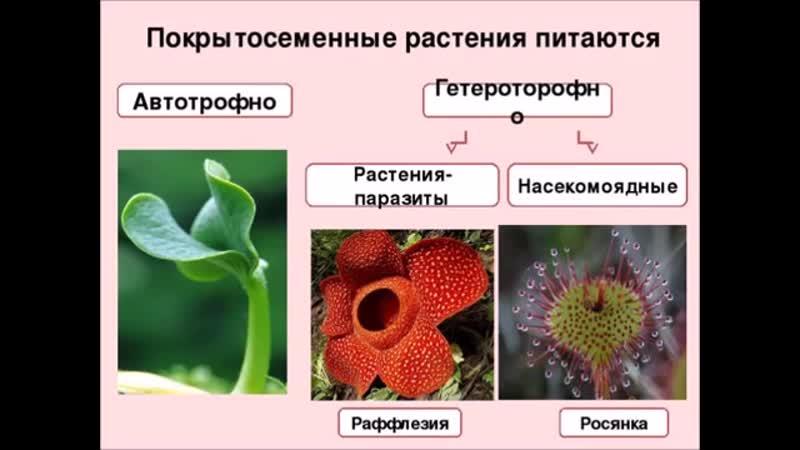
Крапива — не просто сорняк. В ней содержится много полезных веществ, поэтому её используют в медицине.
Фото: flickr.com/svklimkin
Покрытосеменные имеют огромное значение в жизни человека. Они:
- служат пищей для человека и домашних животных;
- являются источником лекарственных компонентов для производства медикаментов и косметики;
- используются как сырье для промышленности;
- просто радуют нас своим прекрасным и разнообразным внешним видом.
Цветковые растения, или покрытосеменные: представители, классификация, размножение
Каждый ученик средней школы слышал про покрытосеменные растения. Неудивительно, ведь им посвящен один из важнейших разделов ботаники. К тому же представители покрытосеменных буквально окружают нас, встречаясь на каждом шагу.
Что такое покрытосеменные растения?
Подавляющее большинство растений, которые видит современный человек каждый день, можно отнести именно к этому отделу. Причем сюда входят цветы, лиственные деревья, кустарники, трава и многое другое. Да, несмотря на то что они различаются сроком жизни, размерами и степенью полезностью, все они представители покрытосеменных, так как объединены рядом важных признаков, о которых расскажем чуть ниже.
Причем сюда входят цветы, лиственные деревья, кустарники, трава и многое другое. Да, несмотря на то что они различаются сроком жизни, размерами и степенью полезностью, все они представители покрытосеменных, так как объединены рядом важных признаков, о которых расскажем чуть ниже.
Но для начала узнаем, когда же впервые возникли эти растения на нашей планете и благодаря чему сумели добиться доминирования.
Когда они появились на Земле
Как удалось установить экспертам, первые представители покрытосеменных цвели еще в меловом периоде – около 140 миллионов лет назад. Так что они являются современниками динозавров, которых успешно пережили. Конечно, за это время растения значительно видоизменились – многие виды погибли в ходе эволюции, а другие – в ходе жизнедеятельности человека. Но это вовсе не помешало им оставаться самыми распространенными представителями растений. Благодаря чему?
В первую очередь свою роль сыграла скорость развития семян. Например, у голосемянных на это уходит несколько лет, в то время как покрытосеменные формируют цветы, которые опыляются и впоследствии дают семена за считаные месяцы.
К тому же они сумели поставить себе на службу ветер, насекомых и даже мелких птиц – все они занимаются опылением, позволяя растениям активно скрещиваться, добиваясь максимального генетического разнообразия, что обеспечивает высокий уровень выживаемости.
Наконец, именно конкуренция между видами расставила каждое из растений в определенную нишу, которую они занимают по сей день.
Какими признаками обладают эти растения
Любому человеку, интересующемуся ботаникой, будет полезно знать основные признаки покрытосеменных растений, чтобы легко опознать их с первого же взгляда. Их довольно много, поэтому перечислим лишь самые основные, понятные и интересные большинству обывателей, а не только узкоспециализированным ученым:
- Наличие цветка – он может быть заметным, красивым и броским или же различимым только под микроскопом. Но именно этот признак объединяет сотни тысяч растений в один отдел.
- Опыление является одним из важнейших этапов жизни растения.
 Выполняться оно может различными способами – как ветром, птицами, водой или насекомыми, так и самостоятельно, без привлечения посторонних организмов.
Выполняться оно может различными способами – как ветром, птицами, водой или насекомыми, так и самостоятельно, без привлечения посторонних организмов. - Семена содержат в себе питательные вещества, которые обеспечивают питание молодого ростка в первые дни или недели жизни – до того, как корневая система станет достаточно мощной, чтобы справиться с этой задачей самостоятельно.
Конечно, существуют и прочие признаки покрытосеменных растений – наличие женских и мужских заростков, способов оплодотворения, триплоидность клеток эндосперма и ряд прочих. Но чтобы разобраться в таких тонкостях, нужно обладать серьезными знаниями по ботанике.
На какие классы делятся
Любой крупный отдел растений делится на соответствующие классы. Конечно, покрытосеменные не являются исключением. Эксперты выделяют здесь классы двудольные и однодольные. Чем они отличаются и как их опознать? Расскажем и об этом.
По внешним признакам растения определить, к какому классу оно относится, довольно сложно – нужно знать ряд свойств и особенностей, а также помнить о многочисленных исключениях, которые здорово усложняют классификацию. Гораздо проще посмотреть именно на семя, из которого выросло растение.
Гораздо проще посмотреть именно на семя, из которого выросло растение.
Например, покрытосеменные класса однодольные имеют цельные семена, не разделенные напополам. Именно сюда относится большинство трав – живут они недолго, зато быстро размножаются, за считаные недели достигая максимального размера. Корневая система мочковатая, но не слишком долговечная, располагающаяся неглубоко от поверхности земли. Цветки в большинстве случаев имеют количество лепестков, кратное трем или, значительно реже, четырем. Но никогда не бывает, чтобы их число делилось без остатка на пять.
Совсем другое дело – класс двудольные. Семя у них, как понятно, разделено на две части и имеет небольшой зародышевый корешок. Корневая система стержневая – более прочная, способная проникать в глубину на несколько метров. Сюда можно отнести не только многие виды трав, но и большинство деревьев, кустарников. Обратите внимание на цветок – он должен иметь количество лепестков, делящееся на четыре или пять без остатка.
Как происходит их размножение
Еще один важный момент – размножение покрытосеменных растений.
Как уже говорилось выше, они используют пыльцу, которая может переноситься с цветка на цветок различными способами: ветром, водой, насекомыми или птицами. Встречаются и самоопыляющиеся, у которых опыление происходит прямо внутри цветка до того, как он раскрылся. Но их количество сравнительно невелико.
По способу опыления их можно разделить на однодомные и двудомные. Первые могут размножаться, даже если растут отдельно от других растений своего вида. Дело в том, что их цветки имеют как тычиночные, так и пестичные цветки. Вторым же для успешного продления рода нужны другие представители своего вида. Это происходит из-за того, что мужские особи имеют только тычиночные цветки, а женские – исключительно пестичные.
На тычинках образуется пыльца, которая должна попасть на пестики. В подавляющем большинстве случаев это происходит биотическим способом: самоопыление, перенос пыльцы птицами или насекомыми. Сюда относится около 80 % покрытосеменных растений. Еще около 19 % опыляется ветром – преимущественно злаки.
Сюда относится около 80 % покрытосеменных растений. Еще около 19 % опыляется ветром – преимущественно злаки.
Созревшие семена часто заключаются в мягкую и вкусную оболочку — плоды, привлекающие не только людей, но и птиц, диких животных. Поедая плоды вместе с семенами, все они становятся носителями и способствуют быстрому распространению растений.
Теперь вы знаете про то, как происходит размножение покрытосеменных растений.
Как правильно – покрытосеменные или цветковые?
Довольно часто люди задаются вопросом о том, как правильно говорить: цветковые растения или покрытосеменные? А также в чем заключается разница между ними?
На самом деле никакой разницы здесь нет. И первое, и второе название обозначают один и тот же отдел растений. К тому же иногда можно услышать третье название – скрытносемянные. Но на сегодняшний день оно считается устаревшим и практически не используется.
Заключение
Как видите, представители покрытосеменных действительно многочисленны. И человечество обязано им действительно многим: начиная с еды и чистого воздуха, заканчивая древесиной и прекрасным настроением при виде роскошных цветов.
И человечество обязано им действительно многим: начиная с еды и чистого воздуха, заканчивая древесиной и прекрасным настроением при виде роскошных цветов.
Особенности покрытосеменных (Реферат) — TopRef.ru
Отличительные особенности покрытосеменных
Наиболее характерной отличительной
особенностью покрытосеменных является
наличие у них своеобразного органа —
цветка, который отсутствует у представителей
других отделов растений. Поэтому
покрытосеменные и называются чаще еще
цветковыми растениями. Семяпочка у них
скрытая, она развивается внутри пестика,
в его завязи, поэтому покрытосеменные
называются иначе пестичными. Пыльца у
покрытосеменных улавливается не
семяпочками, как у голосеменных, а особым
образованием — рыльцем, которым
заканчивается пестик.
Пыльца у
покрытосеменных улавливается не
семяпочками, как у голосеменных, а особым
образованием — рыльцем, которым
заканчивается пестик.
После оплодотворения яйцеклетки из семяпочки образуется семя, а завязь разрастается в плод. Следовательно, семена у покрытосеменных развиваются в плодах, поэтому этот отдел растений и называется покрытосеменные.
Покрытосеменные (Angiospermae), или цветковые (Magnoliophyta) – отдел наиболее совершенных высших растений, имеющих цветок. Ранее включались в отдел семенных растений вместе с голосеменными. В отличие от последних семязачатки цветковых заключены в завязь, образованную сросшимися плодолистиками.
Цветок является генеративным
органом покрытосеменных растений. Он
состоит из цветоножки и цветоложа. На
последнем располагаются околоцветник
(простой или двойной), андроцей
(совокупность тычинок) и гинецей
(совокупность плодолистиков). Каждая
тычинка состоит из тонкой тычиночной
нити и расширенного пыльника, в котором
созревают спермии. Плодолистик цветковых
растений представлен пестиком, который
состоит из массивной завязи и длинного
столбика, вершинная расширенная часть
которого называется рыльце.
Плодолистик цветковых
растений представлен пестиком, который
состоит из массивной завязи и длинного
столбика, вершинная расширенная часть
которого называется рыльце.
Среди покрытосеменных имеются вечнозелёные и листопадные деревья, кустарники и полукустарники, однолетние и многолетние травы. Встречаются эпифиты, использующие в качестве опоры другие растения, паразиты (например, омела), высасывающие воду и органические вещества из других растений, плотоядные растения (росянка, венерина мухоловка), ловящие и переваривающие мелких насекомых.
Покрытосеменные имеют вегетативные органы, обеспечивающие механическую опору, транспорт, фотосинтез, газообмен, а также запасание питательных веществ, и генеративные органы, участвующие в половом размножении. Внутреннее строение тканей наиболее сложно из всех растений; ситовидные элементы флоэмы окружены клетками-спутницами; почти все представители покрытосеменных имеют сосуды ксилемы.
Содержащиеся внутри пыльцевых
зёрен мужские гаметы попадают на рыльце
и прорастают.
Первые цветковые растения
появились в начале мелового периода
около 135 миллионов лет назад (или даже
в конце юрского периода). Вопрос о предке
покрытосеменных в настоящее время
остаётся открытым; наиболее близки к
ним вымершие беннеттитовые, однако,
более вероятно, что вместе с беннеттитами
покрытосеменные обособились от одной
из групп семенных папоротников. Первые
цветковые растения были, по-видимому,
вечнозелёными деревьями с примитивными
цветками, лишёнными лепестков; ксилема
у них всё не имела сосудов.
В середине мелового периода всего за несколько миллионов лет происходит завоевание покрытосеменными суши. Одним из важнейших условий быстрого распространения покрытосеменных была их необычайно высокая эволюционная пластичность. В результате адаптивной радиации, обусловленной экологическими и генетическими факторами (в частности, анеуполидией и полиплоидизацией), образовалось огромное количество различных видов покрытосеменных, входящих в самые разные экосистемы. К середине мелового периода образовалось большинство современных семейств. С цветковыми растениями тесно связана эволюция наземных млекопитающих, птиц и, особенно, насекомых. Последние играют исключительно важную роль в эволюции цветка, осуществляя опыление: яркая окраска, аромат, съедобная пыльца или нектар – всё это средства привлечения насекомых.
Цветковые растения распространены
по всему миру, от Арктики до Антарктики.
В основе их систематики лежит строение
цветка и соцветия, пыльцевых зёрен,
семени, анатомия ксилемы и флоэмы. Почти
250 тысяч видов покрытосеменных делятся
на два класса: двудольные и однодольные,
различающиеся, прежде всего, по количеству
семядолей в зародышах, строению листа
и цветка.
Почти
250 тысяч видов покрытосеменных делятся
на два класса: двудольные и однодольные,
различающиеся, прежде всего, по количеству
семядолей в зародышах, строению листа
и цветка.
Цветковые растения являются одним из ключевых компонентов биосферы: они производят органические вещества, связывают углекислоту и выделяют в атмосферу молекулярный кислород, с них начинаются большинство пастбищных цепей питания. Многие цветковые растения используются человеком для приготовления пищи, строительства жилища, изготовления различных хозяйственных материалов, в медицинских целях.
Покрытосеменные – самый крупный
тип растений, к которому относится более
половины всех известных видов, —
характеризуются рядом четких, резко
отграничивающих их признаков. Наиболее
характерно для них наличие пестика,
образованного одним или несколькими
плодолистиками (макро- и мегаспорофиллами),
сросшимися своими краями, так что в
нижней части пестика образуется замкнутое
полое вместилище – завязь, в которой
развиваются семяпочки (макро- и
мегаспорангии). После оплодотворения
завязь разрастается в плод, внутри
которого находятся развившиеся из
семяпочек семена (или одно семя). Кроме
того для покрытосеменных характерны:
восьмиядерный, или производный из него,
зародышевый мешок, двойное оплодотворение,
триплойдный эндосперм, образующийся
только после оплодотворения, рыльце у
пестика, улавливающее пыльцу, и для
подавляющего большинства – более или
менее типичный цветок с околоцветником.
Из анатомических признаков для
покрытосеменных характерно наличие
настоящих сосудов (трахей), тогда как у
голосеменных развиты только трахеиты,
а сосуды встречаются крайне редко.
После оплодотворения
завязь разрастается в плод, внутри
которого находятся развившиеся из
семяпочек семена (или одно семя). Кроме
того для покрытосеменных характерны:
восьмиядерный, или производный из него,
зародышевый мешок, двойное оплодотворение,
триплойдный эндосперм, образующийся
только после оплодотворения, рыльце у
пестика, улавливающее пыльцу, и для
подавляющего большинства – более или
менее типичный цветок с околоцветником.
Из анатомических признаков для
покрытосеменных характерно наличие
настоящих сосудов (трахей), тогда как у
голосеменных развиты только трахеиты,
а сосуды встречаются крайне редко.
Ввиду большого количества общих
признаков нужно предположить
монофилетическое происхождение
покрытосеменных от какой-то более
примитивной группы голосеменных.
Наиболее ранние и очень отрывочные
ископаемые остатки покрытосеменных
(пыльца, древесина) известны из юрского
геологического периода. Из нижнемеловых
отложений известны тоже немногочисленные
достоверные остатки покрытосеменных,
а в отложениях середины мелового периода
они встречаются сразу в больших
количествах и в значительном разнообразии
форм, которые все принадлежат ко многим
различным ныне живущим семействам и
даже родам.
В качестве предполагаемых предков покрытосеменных указывали различные группы нижестоящих в системе растений – кейтониевые, семенные папоротники, беннеттиты, гнетомые. У кейтониевых были завязь, рыльце, но у них завязь формировалась иначе, чем у покрытосеменных; у них не было даже подобия цветков, спорофиллы их простые и, вероятно, они представляют слепую ветвь эволюции. У беннеттитов были обоеполые своеобразные «цветки», но не было пестиков, и семена их были лишь скрыты между бесплодными чешуями, а не находились внутри плодов, образованных мегаспорофиллами. У семенных папоротников не было цветков, не было покрытосемянности.
Теория происхождения покрытосеменных
из гнетомых предполагает, что наиболее
примитивные покрытосеменные имели
мелкие однополые цветки без околоцветника
или с невзрачным околоцветником. Но по
ряду соображений в настоящее время
более примитивными цветками считают
крупные, обоеполые цветки. Поэтому можно
предположить, что предками современных
покрытосеменных были какие-то вымершие,
очень примитивные голосеменные с
обоеполыми цветками типа шишки
(стробилами), в которых на длинном
цветоложе (оси) были спирально расположены
свободные (несросшиеся друг с другом)
листочки однородного околоцветника,
микроспорофиллы (тычинки) и мегаспорофиллы
(плодолистики). В системе голосеменных
эта группа должна была стоять где-то
между семенными папоротниками и уже
более специализированными беннеттитами
и саговниками.
В системе голосеменных
эта группа должна была стоять где-то
между семенными папоротниками и уже
более специализированными беннеттитами
и саговниками.
Покрытосемянность несомненно
представляла большое преимущество в
смысле защиты семяпочек и развивающихся
семян от всяких неблагоприятных внешних
воздействий и в первую очередь от сухости
воздуха. Но одной покрытосемянностью
все же трудно объяснить быстрое мощное
развитие покрытосеменных и вытеснение
ими господствовавших ранее на земле
архегониальных растений. Русский ботаник
М.И. Голенкин высказал (в 1927г) интересную
гипотезу о причинах победы покрытосеменных
в борьбе за существование. Он предполагает,
что в середине мелового периода по
каким-то общим космогоническим причинам
по всей Земле произошло резкое изменение
освещения и влажности воздуха. Густые
облака, окутывавшие ранее постоянно
Землю, рассеялись и дали доступ ярким
солнечным лучам, в связи с чем, резко
увеличилась сухость воздуха. Громадное
большинство высших архегониальных
растений того времени, не приспособленных
и не сумевших приспособиться к яркому
освещению и сухости воздуха, начало
вымирать или резко сократило области
своего распространения (кроме хвойных,
наиболее ксерофитных).
Покрытосеменные — обзор | ScienceDirect Topics
Покрытосеменные
Покрытосеменные (цветковые растения) содержат 257 000 видов и составляют большую часть разнообразия зеленых, наземных и семенных растений. Эта монофилетическая группа подверглась значительной радиации, начиная с раннего мелового периода (Crane et al. ., 1995; Doyle and Donoghue, 1993). Самые старые окаменелости покрытосеменных представляют собой пыльцевые зерна возрастом 135 млн лет назад. У покрытосеменных появились некоторые новые и важные признаки, которые, вероятно, оказали глубокое влияние на разнообразие цветковых растений: семена, покрытые плодолистиком, редуцированный женский гаметофит, двойное оплодотворение и наличие сосудов как водопроводной системы.Однако у первых покрытосеменных сосудов не было, о чем свидетельствует присутствие трахеид в базальном покрытосеменном Amborella (Field et al. ., 2000). Покрытосеменные не получили широкого распространения до раннего третичного периода, когда произошла радиация современных птиц и млекопитающих; эволюция крупных красочных плодов и семян связана с эволюцией этих групп.
Лигнины покрытосеменных состоят в основном из гваяцильных и сирингильных звеньев с незначительной долей p -гидроксифенильных звеньев.Таким образом, покрытосеменные растения, излучение которых произошло после дивергенции между покрытосеменными и голосеменными, всегда считались единственной филогенетической группой, содержащей сирингиллигнин. Тем не менее, в последние годы многие отчеты, казалось, подвергали сомнению эту давнюю точку зрения. Эти данные свидетельствуют о том, что сирингил лигнины появлялись по крайней мере пять раз в истории растений в результате конвергентной эволюции (у водорослей, мохообразных, плауновидных, голосеменных и покрытосеменных) или что сирингил лигнины являются древним признаком, появившимся у водорослей еще до колонизации суши и утерянным. a posteriori , за исключением некоторых специфических групп и покрытосеменных.Недавние данные, по-видимому, указывают на конвергентную эволюцию, которая предполагает, что сирингил лигнины играют ключевую роль в физиологии растений, даже несмотря на то, что точные функции S-лигнина в растении не выяснены. У покрытосеменных большая часть сирингил-лигнинов расположена в волокнах сосудов, тогда как ксилема в основном состоит из гваяцил-лигнина, что свидетельствует о сильном избирательном давлении в сторону присутствия G-лигнина в проводящих тканях (Peter and Neale, 2004). Аналогичным образом, ксилема Selaginella содержит гваяциллигнин, в то время как сирингил лигнин ограничен эпидермальными и субэпидермальными/кортикальными тканями (рис.6). Аналогичная закономерность была обнаружена и у некоторых папоротников, таких как Ceratopteris (рис. 7). Некоторые сообщения связывают индукцию биосинтеза S-лигнина с реакцией на патогены (Wuyts et al. ., 2006), в то время как другие авторы описывают увеличение количества лигнина после воздействия патогенов, но за счет увеличения единиц H и G. в то время как единицы S остаются неизменными (Gayoso et al. ., 2010; Pomar et al. ., 2004). Структурные различия между G- и S-лигнином обусловлены наличием метоксильной группы в положении 5, в результате чего S-лигнин является более линейным и менее конденсированным.
У покрытосеменных большая часть сирингил-лигнинов расположена в волокнах сосудов, тогда как ксилема в основном состоит из гваяцил-лигнина, что свидетельствует о сильном избирательном давлении в сторону присутствия G-лигнина в проводящих тканях (Peter and Neale, 2004). Аналогичным образом, ксилема Selaginella содержит гваяциллигнин, в то время как сирингил лигнин ограничен эпидермальными и субэпидермальными/кортикальными тканями (рис.6). Аналогичная закономерность была обнаружена и у некоторых папоротников, таких как Ceratopteris (рис. 7). Некоторые сообщения связывают индукцию биосинтеза S-лигнина с реакцией на патогены (Wuyts et al. ., 2006), в то время как другие авторы описывают увеличение количества лигнина после воздействия патогенов, но за счет увеличения единиц H и G. в то время как единицы S остаются неизменными (Gayoso et al. ., 2010; Pomar et al. ., 2004). Структурные различия между G- и S-лигнином обусловлены наличием метоксильной группы в положении 5, в результате чего S-лигнин является более линейным и менее конденсированным. Бонавиц и Чаппл (2010) предположили, что лигнин S придает растениям гибкость. Гибкий полимер может быть важен для травянистых растений, которые каждый год наращивают свою надземную биомассу и, следовательно, должны расти быстро, чего нет у древесных покрытосеменных и голосеменных растений.
Бонавиц и Чаппл (2010) предположили, что лигнин S придает растениям гибкость. Гибкий полимер может быть важен для травянистых растений, которые каждый год наращивают свою надземную биомассу и, следовательно, должны расти быстро, чего нет у древесных покрытосеменных и голосеменных растений.
Известно, что некоторые лигнины ацетилированы по γ-углероду боковой цепи. Недавно сообщалось, что это ацетилирование происходит естественным образом в лигнинах в большей степени, чем считалось ранее (del Río et al ., 2007; Ральф, 1996). Ацетилированный лигнин образуется не в результате ацетилирования растущего полимера лигнина, а из самих ацетилированных монолигнолов, которые включаются в лигнин в результате типичного процесса радикальной полимеризации (Lu and Ralph, 2002, 2008). дель Рио и др. . (2007) сообщили о наличии природного ацетилированного лигнина у всех исследованных покрытосеменных, как древесных, так и травянистых, но не обнаружили его ни у одного из двух голосеменных (сосна и ель). Присутствие ацетилирования было ограничено γ-углеродом и происходило преимущественно на единицах S, тогда как ацетилированные единицы G были едва обнаружены, хотя бамбук и эвкалипт были некоторыми исключениями (del Río et al ., 2007). Таким образом, ацетилирование лигнина было ограничено покрытосеменными растениями и видами с высоким соотношением S/G, о чем свидетельствуют проявления у абаки, кенафа и сизаля (del Río et al ., 2004). Лу и Ральф (2002, 2008) установили, что γ-ацетилированные монолигнолы изменяют структуру лигнина, поскольку группа γ-ОН участвует в некоторых реакциях пост-сочетания, таких как реароматизация хинонметида после β-β’-сочетания и, если γ-ОН группа ацетилирована, эта реакция больше невозможна, что приводит к образованию других новых β-β’-связанных звеньев в полимере.Физиологическая роль ацетилированных лигнинов остается неясной, хотя предполагается их участие в засухоустойчивости, поскольку полимер ацетилированного лигнина является более гидрофобным, чем обычный лигнин, что может сделать сосудистые ткани более гидрофобными, уменьшая потерю воды.
Присутствие ацетилирования было ограничено γ-углеродом и происходило преимущественно на единицах S, тогда как ацетилированные единицы G были едва обнаружены, хотя бамбук и эвкалипт были некоторыми исключениями (del Río et al ., 2007). Таким образом, ацетилирование лигнина было ограничено покрытосеменными растениями и видами с высоким соотношением S/G, о чем свидетельствуют проявления у абаки, кенафа и сизаля (del Río et al ., 2004). Лу и Ральф (2002, 2008) установили, что γ-ацетилированные монолигнолы изменяют структуру лигнина, поскольку группа γ-ОН участвует в некоторых реакциях пост-сочетания, таких как реароматизация хинонметида после β-β’-сочетания и, если γ-ОН группа ацетилирована, эта реакция больше невозможна, что приводит к образованию других новых β-β’-связанных звеньев в полимере.Физиологическая роль ацетилированных лигнинов остается неясной, хотя предполагается их участие в засухоустойчивости, поскольку полимер ацетилированного лигнина является более гидрофобным, чем обычный лигнин, что может сделать сосудистые ткани более гидрофобными, уменьшая потерю воды. Эта гипотеза подтверждается наличием ацетилированного лигнина в некоторых суккулентных агавах (del Río et al. ., 2007). Однако последняя роль не получила широкого признания, поскольку существуют растения с высоким содержанием ацетилированного лигнина, которые не устойчивы к засухе.
Эта гипотеза подтверждается наличием ацетилированного лигнина в некоторых суккулентных агавах (del Río et al. ., 2007). Однако последняя роль не получила широкого признания, поскольку существуют растения с высоким содержанием ацетилированного лигнина, которые не устойчивы к засухе.
Филогенная группа покрытосеменных – обзор
I.B.2. Каллоза и ее роль во время цитокинеза
В то время как роль каллозы SCW во время развития микрогамет остается неясной, меньше неопределенности окружает роль каллозы в межспоровых стенках, где первоначальное разделение отдельных микроспор обеспечивает одну очевидную функцию. Обеспечение впоследствии разлагаемой временной стенки, позволяющей высвобождать микроспоры из тетрады, является возможной второй функцией (например,г. см. Echlin and Godwin, 1968). Однако более важным с точки зрения его влияния на эволюцию покрытосеменных является то, где и когда образуются межспоровые каллозные стенки, потому что это будет определять, сколько пор прорастания будет иметь зрелое пыльцевое зерно.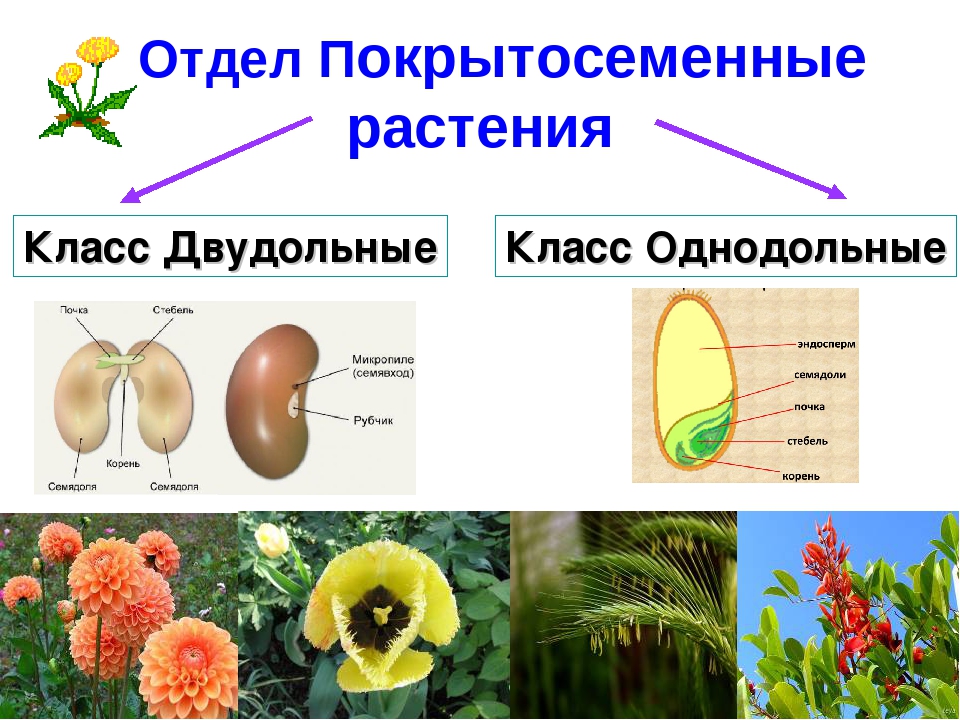 Количество пор, возможно, является наиболее важным признаком в таксономии цветковых растений, поскольку основное деление покрытосеменных растений — это не однодольные и двудольные, а разделение, основанное на типе пыльцы. Базальные покрытосеменные характеризуются однопористой или одноапертурной пыльцой, тогда как более высокопроизводные покрытосеменные (эвдикоты) имеют трехпористую или трехапертурную пыльцу (Группа филогении покрытосеменных, 1998).Трехлопастная пыльца Arabidopsis является примером одного типа трехапертурной пыльцы.
Количество пор, возможно, является наиболее важным признаком в таксономии цветковых растений, поскольку основное деление покрытосеменных растений — это не однодольные и двудольные, а разделение, основанное на типе пыльцы. Базальные покрытосеменные характеризуются однопористой или одноапертурной пыльцой, тогда как более высокопроизводные покрытосеменные (эвдикоты) имеют трехпористую или трехапертурную пыльцу (Группа филогении покрытосеменных, 1998).Трехлопастная пыльца Arabidopsis является примером одного типа трехапертурной пыльцы.
На рис. 4 показаны две общие закономерности появления каллозных стенок после мейоза микроспор, наблюдаемые у цветковых растений (Furness et al., 2002; Furness and Rudall, 2004). Как уже упоминалось, микроспор Arabidopsis претерпевают одновременный цитокинез в конце мейоза II. При одновременном цитокинезе веретена первого и второго мейотических делений взаимодействуют, образуя тетраэдрическую тетраду (рис.4А; Фернесс и др., 2002). В альтернативной модели последовательный цитокинез, стадия диады, соответствующая первому цитоплазматическому делению, происходит после мейоза I. Затем следует второе мейотическое деление. Но поскольку веретена могут принимать различную ориентацию по отношению друг к другу, возникает множество различных расположений тетрады (рис. 4B-E).
В альтернативной модели последовательный цитокинез, стадия диады, соответствующая первому цитоплазматическому делению, происходит после мейоза I. Затем следует второе мейотическое деление. Но поскольку веретена могут принимать различную ориентацию по отношению друг к другу, возникает множество различных расположений тетрады (рис. 4B-E).
Рис. 4. Схема развития общих тетрадных типов. (А) Одновременное деление. (B–E) Последовательное деление. Стрелки указывают на экваториальную плоскость веретена; сплошной кружок обозначает ядро; сплошной кружок внутри маленького круга указывает на ядро, расположенное непосредственно под предыдущим.
(Воспроизведено с разрешения University of Chicago Press из Furness et al., 2002). однодольные растения, хорошо известным примером которых являются злаки (Poaceae) (группа филогении покрытосеменных растений, 1998). У этих растений наблюдаются как одновременные, так и последовательные формы цитокинеза, а также ряд модифицированных вариантов, когда, например, эфемерная клеточная пластинка формируется после мейоза I и впоследствии рассредоточивается, а одновременный цитокинез происходит после мейоза II (Furness et al. , 2002). У пальм (Arecaceae) одновременный и последовательный типы цитокинеза могут происходить даже в пределах одной и той же тычинки (Sannier et al., 2006). Независимо от типа цитокинеза, большинство базальных покрытосеменных производят пыльцу с одной порой на одном полюсе или с узором отверстий, основанным на таком расположении.
, 2002). У пальм (Arecaceae) одновременный и последовательный типы цитокинеза могут происходить даже в пределах одной и той же тычинки (Sannier et al., 2006). Независимо от типа цитокинеза, большинство базальных покрытосеменных производят пыльцу с одной порой на одном полюсе или с узором отверстий, основанным на таком расположении. У более высокопроизводных эвдикотов, группы, которая включает примерно 75% современных видов и большинство растений, традиционно классифицируемых как двудольные (например, Arabidopsis ), микроспоры образуются в результате одновременного цитокинеза, а пыльцевые зерна имеют треугольную или имеют сходный рисунок апертуры (Furness and Rudall, 2004; Angiosperm Phylogeny Group, 1998).Те базальные покрытосеменные растения, которые имеют трехапертурную пыльцу, например члены семейства Illiciaceae, такие как звездчатый анис, также имеют одновременный цитокинез, как и у Arabidopsis (Sampson, 2000; Furness et al., 2002). Таким образом, оказывается, что трехапертурные пыльцевые зерна могут образовываться только в результате одновременного цитокинеза, тогда как одноапертурные пыльцевые зерна могут образовываться либо в результате одновременного, либо последовательного цитокинеза.
На чем основана эта зависимость между образованием каллозной стенки и количеством пор прорастания? Пыльцевые апертуры впервые появляются в онтогенезе экзинового слоя пыльцевой стенки, но их расположение определяется геометрией тетрады и контролируется мейотическим веретеном (Sheldon, Dickinson, 1986; Ressayre et al., 1998, 2005). Комбинации различных вариабельных элементов во время мейоза могут объяснить большинство широко распространенных паттернов. Эти элементы включают время цитокинеза (последовательный, одновременный или промежуточный), ориентацию мейотических осей (тетраэдрическая, тетрагональная, перекрестная, линейная или Т-образная; рис. 4) и способ отложения каллозы для формирования межспоровых стенок. (который может быть либо центростремительным, как у Arabidopsis , либо центробежным, как у большинства однодольных (Nadot et al., 2006).Как правило, апертуры образуются в последних точках цитоплазматического контакта между продуктами мейоза, причем полярные апертуры дополнительно определяются положением полюса веретена во втором мейозе (Ressayre et al.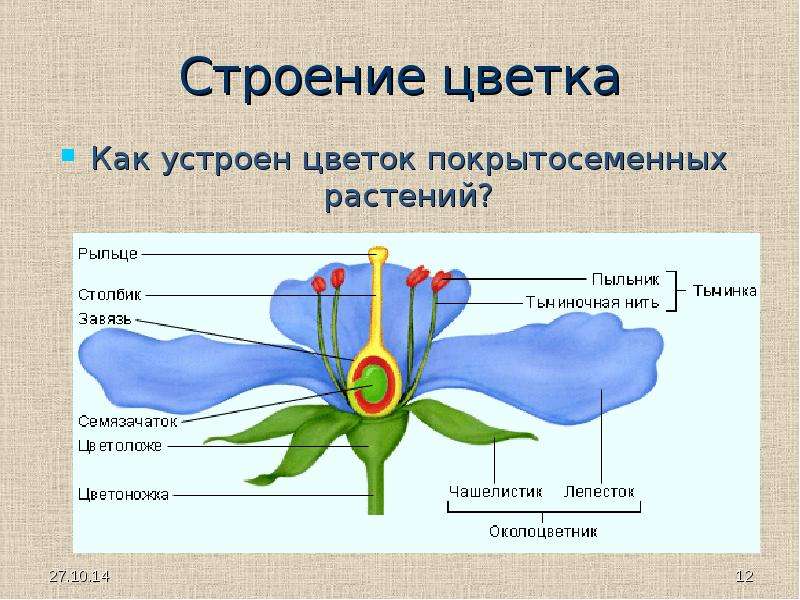 , 2005). Например, в тетраде эвдикота 12 апертур (4 микроспоры × 3 апертуры на микроспору) формируются парами в средних точках (экваториально) по краям тетраэдра, образованного шестью биполярными веретенами (рис. 4А). Места, где впоследствии образуются апертуры, изначально отмечены участками микротрубочек и эндомембран, которые, по-видимому, участвуют в блокировании синтеза матрицы примексина в этих местах (Dickinson and Sheldon, 1984; Munсoz et al., 1995; Рессейр и др., 2002).
, 2005). Например, в тетраде эвдикота 12 апертур (4 микроспоры × 3 апертуры на микроспору) формируются парами в средних точках (экваториально) по краям тетраэдра, образованного шестью биполярными веретенами (рис. 4А). Места, где впоследствии образуются апертуры, изначально отмечены участками микротрубочек и эндомембран, которые, по-видимому, участвуют в блокировании синтеза матрицы примексина в этих местах (Dickinson and Sheldon, 1984; Munсoz et al., 1995; Рессейр и др., 2002).
Большой интерес вызывают гены и белки, участвующие в мейотическом цитокинезе самцов. По самой своей природе мутации, влияющие на функцию этих генов, также могут снижать жизнеспособность пыльцы, поэтому для их идентификации широко используются генетические скрининги мужской стерильности (см., например, Caryl et al., 2003; Johnson-Brousseau and McCormick, 2004).
Как и ожидалось, некоторые мутации мужской стерильности специфически влияют на цитокинез и связаны с образованием или растворением каллозы. У Arabidopsis гены GLS , AtGSL1 и AtGSL5 , по-видимому, играют важную и перекрывающуюся роль в синтезе межспоровых стенок (Enns et al., 2005). Развитие микрогамет у мутантных растений atgsl1 или atgsl5 протекает нормально до завершения мейоза I (т.е. двухклеточный микроспороцит, окруженный мозолистым SCW), что позволяет предположить, что GSL1 и GSL5 не требуются на этих ранних стадиях. Однако в то время как растения, гомозиготные по gsl1 , производят нормальную пыльцу, 15 % пыльцы от gsl5 -гомозиготных растений и 30 % пыльцы от gsl1 /+ gsl5 / gls5 растений (т.е. растения, гомозиготные по мутации gsl1 и гетерозиготные по мутации gsl5 ) и gsl1 / gsl1 gsl5 /+, растения маленькие, сморщенные, с неуместными и деформированными устьями. Поскольку эти пыльцевые зерна завершили мейоз, это указывает на то, что дефицит GSL1 и GSL5 влияет на более поздние стадии развития пыльцы, когда в норме каллоза отсутствует.
У Arabidopsis гены GLS , AtGSL1 и AtGSL5 , по-видимому, играют важную и перекрывающуюся роль в синтезе межспоровых стенок (Enns et al., 2005). Развитие микрогамет у мутантных растений atgsl1 или atgsl5 протекает нормально до завершения мейоза I (т.е. двухклеточный микроспороцит, окруженный мозолистым SCW), что позволяет предположить, что GSL1 и GSL5 не требуются на этих ранних стадиях. Однако в то время как растения, гомозиготные по gsl1 , производят нормальную пыльцу, 15 % пыльцы от gsl5 -гомозиготных растений и 30 % пыльцы от gsl1 /+ gsl5 / gls5 растений (т.е. растения, гомозиготные по мутации gsl1 и гетерозиготные по мутации gsl5 ) и gsl1 / gsl1 gsl5 /+, растения маленькие, сморщенные, с неуместными и деформированными устьями. Поскольку эти пыльцевые зерна завершили мейоз, это указывает на то, что дефицит GSL1 и GSL5 влияет на более поздние стадии развития пыльцы, когда в норме каллоза отсутствует. Примечательно, что растения gls1 /+ gsl5 / gsl5 также образуют увеличенные многоядерные пыльцевые зерна с более чем тремя порами: отсутствие межспоровых стенок в этих пыльцевых зернах указывает на то, что GSL1 и GSL5 необходимы для синтеза межспоровых стенок. но не мозолистый SCW (Enns et al., 2005). Этот фенотип можно противопоставить эффекту мутаций в ATGSL2 , где нет общего SCW, но цитокинез протекает нормально (Nishikawa et al., 2005).
Примечательно, что растения gls1 /+ gsl5 / gsl5 также образуют увеличенные многоядерные пыльцевые зерна с более чем тремя порами: отсутствие межспоровых стенок в этих пыльцевых зернах указывает на то, что GSL1 и GSL5 необходимы для синтеза межспоровых стенок. но не мозолистый SCW (Enns et al., 2005). Этот фенотип можно противопоставить эффекту мутаций в ATGSL2 , где нет общего SCW, но цитокинез протекает нормально (Nishikawa et al., 2005).
Отсутствие образования межспоровых стенок также наблюдается у растений Arabidopsis , несущих аллельные мутации тетраспоры и шпильки (Hűlskamp et al., 1997; Spielman et al., 1997). Поскольку у этих мутантов не происходит цитокинеза микроспор, образуются большие многоядерные ценоцитарные пыльцевые зерна с аберрантным количеством неуместных апертур (Spielman et al., 1997). В соответствии с предполагаемой ролью микротрубочек в регуляции как расположения плоскостей деления, так и участков образования будущих пор, продукт TETRASPORE представляет собой кинезин, двигательный белок, который связывается с микротрубочками и участвует в переносе везикул (Yang et al. ., 2003).
., 2003).
После завершения цитокинеза разрушение отдельных SCW обычно приводит к высвобождению микроспор. Однако некоторые виды естественным образом выделяют пыльцу в виде тетрады сросшихся пыльцевых зерен.Это наблюдается у представителей семейства горных перцев (Winteraceae), а также у кувшинок (Nymphaeales), камыша (Typhaceae), вересковых (Ericaceae), энотера (Onagraceae) и акаций (Fabaceae) (Smyth, 1994; Копенхейвер и др., 2000). В этих семействах в межспоровых поперечных стенках тетрады часто имеется мало или совсем нет каллозы, что позволяет слоям экзин соседних микроспор сливаться и препятствует отделению пыльцевых зерен (Prakash et al., 1992; Scott et al., 2004). Растения Arabidopsis с одной из трех неаллельных мутаций квартета ( qrt ) ( qrt1-3 ) также производят слитые тетрады пыльцевых зерен (Preuss et al., 1994; Copenhaver et al.; Rhe, 2000). и др., 2003). Хотя дефектная или отсроченная деградация каллозы была предложена в качестве причины, по которой qrt микроспор не могут разделиться (Echlin and Godwin, 1968; Izhar and Frankel, 1971), анализ qrt мутантов предполагает, что вместо этого тетрады удерживаются вместе изменения пектиновых компонентов в первичной клеточной стенке вокруг микроспороцита (Rhee and Somerville, 1998).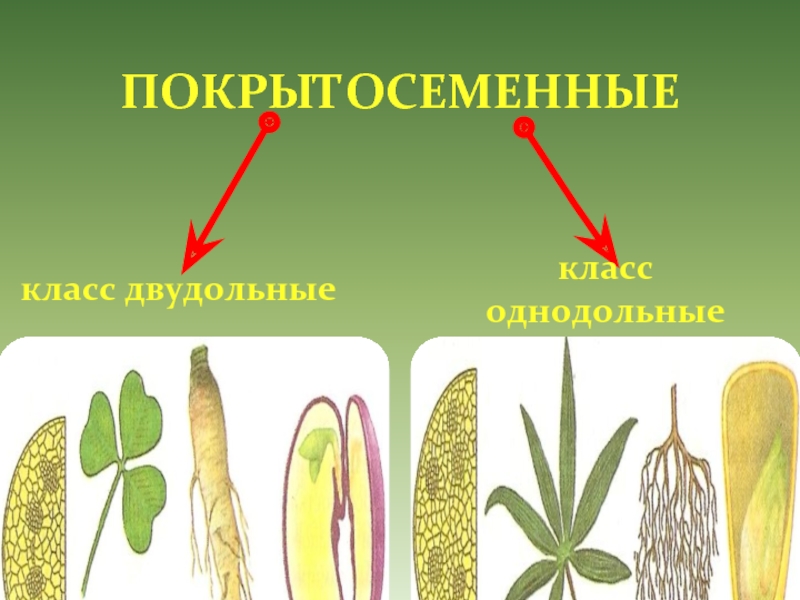 В соответствии с этим недавно было показано, что QRT1 кодирует пектинметилэстеразу, а QRT3 — эндо-полигалактуроназу (Rhee et al., 2003; Francis et al., 2006).
В соответствии с этим недавно было показано, что QRT1 кодирует пектинметилэстеразу, а QRT3 — эндо-полигалактуроназу (Rhee et al., 2003; Francis et al., 2006).
Американский журнал ботаники
Я только что прочитал Эссе Болла. Это довольно смело. Быстрое развитие, насколько мы можем судить, всех высших растений в новейшее геологическое время представляет собой отвратительную тайну. Конечно, было бы большим шагом, если бы мы могли поверить, что высшие растения сначала могли жить только на высоком уровне; но пока экспериментально [доказано] , что цикадовые, папоротники и т. д., могут выдерживать гораздо больше углекислоты, чем высшие растения, эта гипотеза кажется мне слишком опрометчивой. Сапорта полагает, что произошло поразительно быстрое развитие высших растений, как только 90 140 [как] 90 139 насекомых, посещающих цветы, развились и способствовали скрещиванию. Мне бы хотелось, чтобы вся эта проблема была решена. Я воображал, что, может быть, в течение долгих веков в Южном полушарии существовал небольшой изолированный континент, служивший родиной высших растений, — но это жалко бедное предположение 90 140 . — Отрывок из письма, написанного Чарльзом Дарвином 22 июля 1879 г. Джозефу Хукеру
— Отрывок из письма, написанного Чарльзом Дарвином 22 июля 1879 г. Джозефу Хукеру
Очарование и разочарование Чарльза Дарвина эпическими эволюционными событиями, связанными с происхождением и ранним излучением цветковых растений, ходят легенды. Пожалуй, никакая другая группа организмов не заслуживала внимания Дарвина в таких драматических выражениях: «отвратительная тайна», «самое загадочное явление», «ничего… более необычного». Но из всех комментариев Дарвина о ранней истории эволюции цветковых растений его «отвратительная тайна» захватила воображение целых поколений биологов растений.Всего через несколько месяцев после того, как письмо Дарвина Джозефу Хукеру от 22 июля 1879 г. (рис. 1A–G1B1C1D1E1F1G) было впервые опубликовано в 90 139, More Letters of Charles Darwin 90 140 (17), биологи неустанно используют фразу «отвратительная тайна» посредством современного синтеза и к современному синтезу молекулярной филогенетики, генетики развития, морфологии и палеоботаники (например, 55; 29; 59; 45; 54; 62; 1; 58; 47; 12, 3; 3; 6; 39; 18; 22; 23; 26; 60).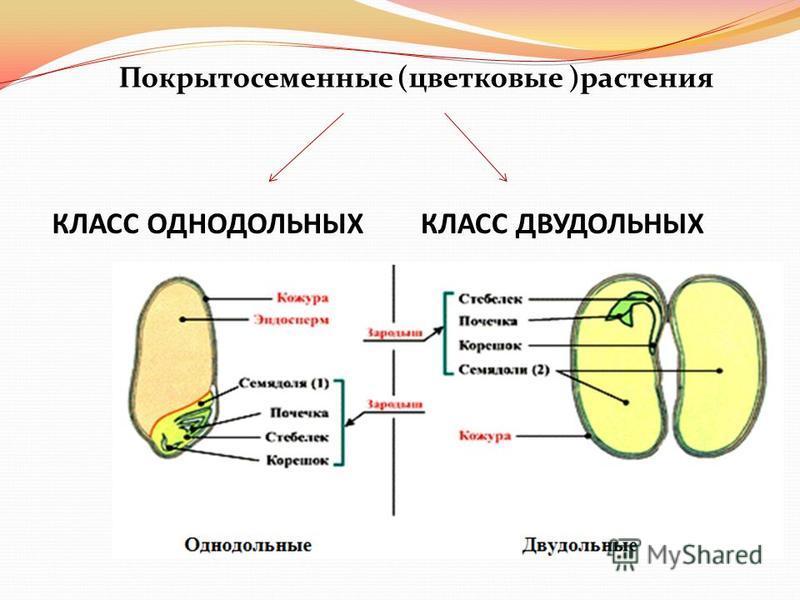 Поиск Google в Интернете по словам «отвратительная тайна» и «Дарвин» даст сотни (если не тысячи) результатов, часто в заголовках научных статей упоминается «загадка разгадана».
Поиск Google в Интернете по словам «отвратительная тайна» и «Дарвин» даст сотни (если не тысячи) результатов, часто в заголовках научных статей упоминается «загадка разгадана».
Письмо Чарльза Дарвина Джозефу Далтону Хукеру, написанное 22 июля 1879 г. (происхождение: Библиотека Кембриджского университета DAR 95: 485–488). В этом письме Дарвин называет раннюю эволюцию цветковых растений «отвратительной тайной». Он также проявляет интерес к идее Гастона де Сапорта о том, что коэволюционный набор взаимодействий между покрытосеменными и насекомыми мог быть центральным элементом быстрого разнообразия цветковых растений в середине мелового периода.Это письмо является прекрасным примером переписки Дарвина с Хукером, наполненной обменом научной информацией и вопросами, последними сведениями о сочинениях и публикациях Дарвина, семейными делами (отпуск) и повседневными делами (проблемы заражения щитовкой на растении, одолженном Дарвину из Кью). Почерк Дарвина часто очень трудно разобрать, но почерк в этом письме на самом деле довольно хороший по сравнению с другими почерками этого позднего периода его жизни (Э. Смит, Darwin Correspondence Project, библиотека Кембриджского университета, личное сообщение).Далее следует транскрипция письма. Обратите внимание, что цифры 485, 486, 487 и 488, написанные карандашом на первой, третьей, пятой и седьмой страницах, являются отметками класса, связанными с архивами библиотеки Кембриджского университета. На первой странице «/79» после даты и аннотация слева «отправлено 23 июля 1979 г.» написаны карандашом и, возможно, были добавлены Фрэнсисом Дарвином, когда он расшифровывал и публиковал части этого письма в 1903 году. На последней странице письма карандашное «Нет», возможно, было добавлено Джозефом Хукером в ответ на запрос Дарвина о возвращении зараженного чешуей растения Smilax в Кью.Цифровые изображения этого письма из коллекции Дарвина, любезно предоставленные библиотекой Кембриджского университета.
Почерк Дарвина часто очень трудно разобрать, но почерк в этом письме на самом деле довольно хороший по сравнению с другими почерками этого позднего периода его жизни (Э. Смит, Darwin Correspondence Project, библиотека Кембриджского университета, личное сообщение).Далее следует транскрипция письма. Обратите внимание, что цифры 485, 486, 487 и 488, написанные карандашом на первой, третьей, пятой и седьмой страницах, являются отметками класса, связанными с архивами библиотеки Кембриджского университета. На первой странице «/79» после даты и аннотация слева «отправлено 23 июля 1979 г.» написаны карандашом и, возможно, были добавлены Фрэнсисом Дарвином, когда он расшифровывал и публиковал части этого письма в 1903 году. На последней странице письма карандашное «Нет», возможно, было добавлено Джозефом Хукером в ответ на запрос Дарвина о возвращении зараженного чешуей растения Smilax в Кью.Цифровые изображения этого письма из коллекции Дарвина, любезно предоставленные библиотекой Кембриджского университета.
Рис. 1А. Письмо Чарльза Дарвина Джозефу Далтону Хукеру, написанное 22 июля 1879 г., стр. 1. Мой дорогой Хукер, если мне не изменяет память, Дайер [Уильям Тернер Тиселтон-Дайер, помощник директора садов Кью] уехал из Кью на каникулы, и поэтому я Напишите вам, чтобы узнать, нет ли у вас случайно семян Lathyrus aphaca или любых молодых саженцев, 2 или 3 из которых можно было бы посадить в горшки.Если я не получу ответа, я пойму, что вы не можете мне помочь. — Я хочу проверить, являются ли усики афелиотропными, поскольку я записал, что они очень мало вращаются, я предполагаю, что они могут найти
Письмо Чарльза Дарвина Джозефу Далтону Хукеру, написанное 22 июля 1879 г., стр. 2. поддержка, наклоняясь к любым темным предметам. — Наша книга о движении растений [Сила движения растений , 1880 г., написанная Чарльзом Дарвином при «содействии» его сына Фрэнсиса Дарвина], я думаю, будет содержать много нового материала, но будет невыносимо скучно. В последнее время я очень много работаю и хочу отдохнуть и переодеться, поэтому мы все едем 1 августа -й в Конистон на месяц. Это ужасное путешествие для меня. — Давненько я не слышал никаких новостей о вас и о вас, о том, что вы делаете и что собираетесь делать
В последнее время я очень много работаю и хочу отдохнуть и переодеться, поэтому мы все едем 1 августа -й в Конистон на месяц. Это ужасное путешествие для меня. — Давненько я не слышал никаких новостей о вас и о вас, о том, что вы делаете и что собираетесь делать
Письмо Чарльза Дарвина Джозефу Далтону Хукеру, написанное 22 июля 1879 г., стр. 3. Сделать. Фрэнк [Фрэнсис] возвращается в начале следующего месяца из Вюрцбурга, где он довольно много работал над различными предметами и практиковался в вскрытии, нарезании кусочков и т. д.Я только что прочитал эссе Болла. Это довольно смело. Быстрое развитие, насколько мы можем судить, всех высших растений в новейшее геологическое время представляет собой отвратительную тайну. Конечно, было бы большим шагом, если бы мы могли поверить, что высшие растения сначала могли жить только на высоком уровне; но пока экспериментально [доказано]
Письмо Чарльза Дарвина Джозефу Далтону Хукеру, написанное 22 июля 1879 года, страница 4. что саговники, папоротники и т. д. могут выдерживать гораздо больше углекислоты, чем высшие растения, эта гипотеза кажется мне слишком опрометчивой. Сапорта полагает, что произошло удивительно быстрое развитие высших растений, как только развились посещающие цветы насекомые и благоприятствовали перекрестному скрещиванию. Мне бы хотелось, чтобы вся эта проблема была решена. Я воображал, что, возможно, в течение долгих веков в Южном полушарии существовал небольшой изолированный континент, который служил местом рождения высших растений; но это
что саговники, папоротники и т. д. могут выдерживать гораздо больше углекислоты, чем высшие растения, эта гипотеза кажется мне слишком опрометчивой. Сапорта полагает, что произошло удивительно быстрое развитие высших растений, как только развились посещающие цветы насекомые и благоприятствовали перекрестному скрещиванию. Мне бы хотелось, чтобы вся эта проблема была решена. Я воображал, что, возможно, в течение долгих веков в Южном полушарии существовал небольшой изолированный континент, который служил местом рождения высших растений; но это
Письмо Чарльза Дарвина Джозефу Далтону Хукеру, написанное 22 июля 1879 года, страница 5.это ужасно бедная догадка. Странно, что Болл не ссылается на тот очевидный факт, что альпийские растения должны были существовать до ледникового периода, многие из которых вернулись бы в горы после ледникового периода, когда климат снова стал теплым. Я всегда так относил себя к горечавкам и т. д.
д.
Письмо Чарльза Дарвина Джозефу Далтону Хукеру, написанное 22 июля 1879 года, страница 6.Боллу следовало также учитывать альпийских насекомых, обычных для арктических регионов. Не знаю, как у вас, но моя вера в Ледниковую миграцию ничуть не поколеблена. Мой дорогой старый друг, искренне ваш Ch. Дарвин
Письмо Чарльза Дарвина Джозефу Далтону Хукеру, написанное 22 июля 1879 г., стр. 7. P.S. Когда мы уедем из дома, мне придется вернуть несколько растений Кью. — Ваше растение Smilax aspera было повреждено щитовкой, обнаруженной совсем недавно.— Стоит ли возвращаться? Это большой куст.
В течение прошлого века ужасная тайна Дарвина стала синонимом сложностей и часто кажущихся неразрешимыми вопросов, связанных с происхождением и самыми ранними фазами истории эволюции покрытосеменных. Значения, приписываемые отвратительной тайне Дарвина, сильно различаются и включают филогенетические отношения покрытосеменных растений с другими линиями семенных растений; филогенетические отношения основных клад внутри покрытосеменных растений; поиск ископаемых предшественников цветковых растений; поиск самых ранних ископаемых цветковых растений; эволюционное происхождение цветка, плодолистика и множество других вопросов гомологии уникальных характеристик покрытосеменных растений; и относительно резкий рост экологического доминирования покрытосеменных растений в меловой период.
Значения, приписываемые отвратительной тайне Дарвина, сильно различаются и включают филогенетические отношения покрытосеменных растений с другими линиями семенных растений; филогенетические отношения основных клад внутри покрытосеменных растений; поиск ископаемых предшественников цветковых растений; поиск самых ранних ископаемых цветковых растений; эволюционное происхождение цветка, плодолистика и множество других вопросов гомологии уникальных характеристик покрытосеменных растений; и относительно резкий рост экологического доминирования покрытосеменных растений в меловой период.
Помимо простого исключения этой двухсловной фразы из объемистых сочинений Дарвина, стоит задаться вопросом, о чем именно размышлял Дарвин (рис. 2), когда писал об «отвратительной тайне». Этот важный вопрос никогда по-настоящему не рассматривался, если не считать того, что Дарвин был озадачен происхождением и ранней историей цветковых растений. Учитывая всеобщую склонность цитировать Дарвина, когда это возможно, кажется вполне уместным, чтобы мысли Чарльза Дарвина об этих основополагающих эволюционных событиях были систематически проанализированы. Как мы увидим, с 1870-х годов и до самого последнего года своей жизни Чарльз Дарвин был глубоко озадачен ранней историей эволюции покрытосеменных, и в отвратительной тайне Дарвина есть нечто большее, чем можно уловить в двух словах. словесная фраза.
Как мы увидим, с 1870-х годов и до самого последнего года своей жизни Чарльз Дарвин был глубоко озадачен ранней историей эволюции покрытосеменных, и в отвратительной тайне Дарвина есть нечто большее, чем можно уловить в двух словах. словесная фраза.
Чарльз Дарвин в 1877 и 1878 годах. Слева . Дарвин в 1878 году, сфотографированный его сыном Леонардом. Справа . Дарвин на своей лошади «Томми» в Даун-Хаусе в конце 1870-х годов.Под копией этой фотографии он написал: «Ура, сегодня писем нет!» Его комментарий является прекрасным напоминанием о чрезвычайной важности высокоэффективной британской почтовой службы для обеспечения того, чтобы его обширная переписка с коллегами по всему миру, в том числе с теми, кто помог сформировать его взгляды на «отвратительную тайну», доставлялась в Даун-Хаус и обратно. . Изображения Дарвина из коллекции Дарвина, предоставлены библиотекой Кембриджского университета.
В чем заключалась ужасная тайна Дарвина?
Особое увлечение и разочарование Дарвина ранней эволюцией покрытосеменных, по-видимому, всерьез проявились за несколько лет до его часто цитируемого письма (22 июля 1879 г. ) Джозефу Хукеру (рис.3). В переписке (8 марта 1875 г.) со швейцарским ботаником, энтомологом и палеонтологом Освальдом Хеером (рис. 3) Дарвин зашел так далеко, что отметил, что «внезапное появление такого большого количества двудольных в верхнем мелу кажется мне весьма озадачивающим». явление всем, кто верит в какую-либо форму эволюции, особенно тем, кто верит в чрезвычайно постепенную эволюцию, против которой, я знаю, вы решительно выступаете» (17, с. 239). Хир был сторонником возможности быстрой (по сути точечной или скачкообразной) эволюции, к большому разочарованию Дарвина.
) Джозефу Хукеру (рис.3). В переписке (8 марта 1875 г.) со швейцарским ботаником, энтомологом и палеонтологом Освальдом Хеером (рис. 3) Дарвин зашел так далеко, что отметил, что «внезапное появление такого большого количества двудольных в верхнем мелу кажется мне весьма озадачивающим». явление всем, кто верит в какую-либо форму эволюции, особенно тем, кто верит в чрезвычайно постепенную эволюцию, против которой, я знаю, вы решительно выступаете» (17, с. 239). Хир был сторонником возможности быстрой (по сути точечной или скачкообразной) эволюции, к большому разочарованию Дарвина.
Ключевые корреспонденты Чарльза Дарвина в его обсуждениях ранней эволюции цветковых растений. Вверху слева . Джозеф Далтон Хукер в 1877 году на перевале Ла-Вета в Колорадо, США. Вверху справа . Освальд Хир (1809–1883), дата неизвестна. Воспроизведено из Oswald Heer: Lebensbild eines schweizerischen Naturforschers (53). Внизу слева . Джон Болл (1818–1889), дата неизвестна.Джон Болл был заядлым натуралистом, который учился у Хенслоу в Кембридже. В качестве заместителя государственного секретаря в Министерстве по делам колоний он продвигал ботанические интересы Королевского ботанического сада в Кью (34 года; 19 лет) и был альпийским экспертом и ботаником. Некролог 34 дает отличный очерк его жизни. Воспроизведено из Сэр Джозеф Далтон Хукер, путешественник и коллекционер растений (19). Внизу справа . Гастон де Сапорта (1823–1895), между 1880 и 1885 годами.
Внизу слева . Джон Болл (1818–1889), дата неизвестна.Джон Болл был заядлым натуралистом, который учился у Хенслоу в Кембридже. В качестве заместителя государственного секретаря в Министерстве по делам колоний он продвигал ботанические интересы Королевского ботанического сада в Кью (34 года; 19 лет) и был альпийским экспертом и ботаником. Некролог 34 дает отличный очерк его жизни. Воспроизведено из Сэр Джозеф Далтон Хукер, путешественник и коллекционер растений (19). Внизу справа . Гастон де Сапорта (1823–1895), между 1880 и 1885 годами.
Летопись окаменелостей ранних покрытосеменных растений, с которой столкнулись Чарльз Дарвин, Освальд Хир и другие (включая Джона Болла и Гастона де Сапорта, как будет показано ниже) в середине 1870-х годов, была богата радиацией покрытосеменных от среднего до позднего мела, но практически не содержала представления о раннемеловых началах эволюции покрытосеменных.Хотя стратиграфическое разрешение в то время было рудиментарным, любой, кто интересовался ранней эволюционной историей цветковых растений, ясно увидел бы контраст между фактическим отсутствием покрытосеменных растений в раннем меловом периоде и их явным восхождением к экологическому и биогеографическому доминированию в позднем меловом периоде.
Говоря драматическим языком, Освальд Хир подробно обсуждал раннюю летопись окаменелостей покрытосеменных с Дарвином в письме от 1 марта 1875 г. (в то время Хир был в разгаре публикации серии из семи томов, Flora Fossilis Arctica , 1868–1883) .Хир писал (происхождение: Библиотека Кембриджского университета, DAR 166: 130): «В Европе также отсутствуют двудольные нижнего мелового периода; однако они встречаются в большом изобилии в верхнем мелу… как в Европе, так и в Америке… [I] возможно, что некоторые из этих видов [двудольных покрытосеменных] еще могут быть обнаружены там [в нижнем мелу Европы и Америки]. Даже в этом случае, если мы говорим, что двудольные начинаются с верхнего мела, мы все же должны признать, что эта часть растительного царства, составляющая основную часть современной растительности, появляется относительно поздно и что в геологическом отношении она претерпела существенную трансформацию. в течение короткого промежутка времени.
Понимание Чарльзом Дарвином летописи окаменелостей, в том виде, в каком она была известна в 1875 году , было абсолютно ясным: ранняя диверсификация и биогеографическое распространение покрытосеменных были удивительно быстрыми. Это резкое и относительно широкое появление почти современного разнообразия покрытосеменных растений в середине мелового периода, практически без предшествующей летописи окаменелостей, поставило Дарвина в крайне неудобное положение. Если бы летопись окаменелостей была точным индикатором прошлых событий, она представляла бы серьезный вызов его общему представлению о градуализме как способе трансформации, хотя Дарвин, безусловно, допускал возможность некоторых быстрых эволюционных изменений (42; 28; 48; 27). .Это, в свою очередь, привело Дарвина к шестилетней одиссее, чтобы объяснить либо чрезвычайно быстрый темп эволюционного разнообразия, либо поразительно длинную и отсутствующую летопись окаменелостей самых ранних (и постепенных) фаз эволюции покрытосеменных. Как заметил Майр (1982, стр. 509), «всю свою жизнь Дарвин прилагал огромные усилия, чтобы реконструировать постепенную эволюцию явлений, которые на первый взгляд казались явно результатом внезапного возникновения».
Как заметил Майр (1982, стр. 509), «всю свою жизнь Дарвин прилагал огромные усилия, чтобы реконструировать постепенную эволюцию явлений, которые на первый взгляд казались явно результатом внезапного возникновения».
Дарвин разгадывает чудовищную тайну
Склонность Дарвина придерживаться градуалистской точки зрения на темпы эволюционных инноваций привела его к постулату, что до мелового периода цветковых растений покрытосеменные растения медленно развивались и диверсифицировались на отдаленном (и более не существующем) массиве суши, возможно, на юге. полушарие.Как следствие, цветковые растения отсутствовали в летописи окаменелостей до середины мелового периода, когда они, наконец, распространились за пределы этой ограниченной территории. Как выразился Дарвин Хееру, «растения этого большого отдела, должно быть, в значительной степени развились в какой-то изолированной области, откуда благодаря географическим изменениям им наконец удалось ускользнуть и быстро распространиться по миру» (письмо к Освальду Хееру, 8). март 1875 г., 17, стр. 240). Четыре года спустя Дарвин снова намекнул на эту идею в своем отвратительном таинственном письме к Хукеру (17, стр.21, 22): «Я воображал, что, возможно, в течение долгих веков в Южном полушарии существовал небольшой изолированный континент, служивший родиной высших растений, — но это ужасно жалкое предположение».
март 1875 г., 17, стр. 240). Четыре года спустя Дарвин снова намекнул на эту идею в своем отвратительном таинственном письме к Хукеру (17, стр.21, 22): «Я воображал, что, возможно, в течение долгих веков в Южном полушарии существовал небольшой изолированный континент, служивший родиной высших растений, — но это ужасно жалкое предположение».
Ближе к концу своей жизни Дарвин вернулся к идее утерянной палеонтологической летописи самых ранних фаз диверсификации покрытосеменных растений (16, стр. 248) в письме Хукеру (6 августа 1881 г.). «Нет ничего более необыкновенного в истории Растительного царства, как мне кажется, чем очень внезапное или резкое развитие высших растений.Иногда я размышлял о том, не существовало ли где-то в течение долгих веков крайне изолированного континента, возможно, вблизи Южного полюса». Акцент Дарвина на слове «очевидно» показывает его глубоко укоренившееся понимание того, что летопись окаменелостей может быть удивительно неполной, а также его постоянный скептицизм в отношении того, что серьезное эволюционное излучение такой величины, которое наблюдалось (по состоянию на 1881 г. ) у покрытосеменных среднего мела, действительно могло быть такой резкий.
) у покрытосеменных среднего мела, действительно могло быть такой резкий.
Менее чем через неделю (письмо от 11 августа 1881 г.; 17, с.26), Дарвин повторил эти же тезисы Хукеру. «Я был настолько поражен кажущимся внезапным появлением высших фанерогам, что мне иногда казалось, что развитие могло медленно продолжаться в течение огромного периода времени на каком-нибудь изолированном континенте или большом острове, возможно, вблизи Южного полюса». Это письмо, кажется, содержит последние записанные слова Дарвина о ранней эволюции покрытосеменных растений.
Как Дарвин написал об ужасной тайне
Отвратительное загадочное письмо Дарвина Хукеру от 22 июля 1879 г. напрямую связано с его сильным желанием смягчить предполагаемую быструю скорость диверсификации ранних покрытосеменных растений.Толчком к написанию этого письма послужило чтение Дарвином эссе «О происхождении флоры европейских Альп», поставленного Джоном Боллом (рис. 3) на заседании Королевского географического общества 9 июня 1879 г. и впоследствии опубликованного в сентябре того же года (2). В этой статье, которая в значительной степени посвящена биогеографическим аспектам альпийских флор, Болл также непосредственно затронул вопрос о ранней эволюционной истории покрытосеменных растений. В письме Дарвину от 8 августа 1879 года Болл отметил, что он организовал отправку препринта этой публикации Дарвину в начале лета.Таким образом, мы можем с уверенностью предположить, что эта рукопись была у Дарвина в руках не позднее июня или июля 1879 г.
3) на заседании Королевского географического общества 9 июня 1879 г. и впоследствии опубликованного в сентябре того же года (2). В этой статье, которая в значительной степени посвящена биогеографическим аспектам альпийских флор, Болл также непосредственно затронул вопрос о ранней эволюционной истории покрытосеменных растений. В письме Дарвину от 8 августа 1879 года Болл отметил, что он организовал отправку препринта этой публикации Дарвину в начале лета.Таким образом, мы можем с уверенностью предположить, что эта рукопись была у Дарвина в руках не позднее июня или июля 1879 г.
Эссе Болла (1879, стр. 579) представляет собой важное окно в летопись окаменелостей покрытосеменных растений, с которой столкнулся Дарвин в конце 1870-х годов. «Появление экзогенных растений высшего типа [двудольных покрытосеменных] не раскрывается прямыми свидетельствами примерно до середины [C]речного периода. Затем вдруг в отложениях, широко распространенных в северном полушарии, мы встречаем множество видов, принадлежащих к самым разным типам, но большей частью настолько похожих на живые растения, что палеонтологи не колеблясь относят многие из них к существующим. роды.Затем Болл (стр. 580) задал вопрос лаконично. «Но если в начале самой ранней из доступных нам глав истории эволюция цветковых растений и особенно экзогенов [двудольных покрытосеменных] уже зашла так далеко, то где, спрошу я, мы должны смотреть? для более ранних форм, предковых типов, от которых произошли наши нынешние группы? И где опять-таки гораздо более отдаленные формы, которые служили мостом через интервал, столь сбивающий с толку ботаника, между эндогенами [однодольными] и экзогенами?»
роды.Затем Болл (стр. 580) задал вопрос лаконично. «Но если в начале самой ранней из доступных нам глав истории эволюция цветковых растений и особенно экзогенов [двудольных покрытосеменных] уже зашла так далеко, то где, спрошу я, мы должны смотреть? для более ранних форм, предковых типов, от которых произошли наши нынешние группы? И где опять-таки гораздо более отдаленные формы, которые служили мостом через интервал, столь сбивающий с толку ботаника, между эндогенами [однодольными] и экзогенами?»
Возможно, невольно Болл уловил суть дарвиновской дилеммы покрытосеменных растений.«На мой взгляд, нет альтернативы между отказом от доктрины эволюции и признанием того, что происхождение существующих видов цветковых растений неизмеримо более отдалено, чем период, относительно которого у нас есть прямые свидетельства. Преодолимая трудность состоит в полном отсутствии таких свидетельств» (2, с. 580). Как четыре года назад Дарвин признался Хиру (и отметил выше), «внезапное появление такого большого количества двудольных в Верхнем Меле кажется мне весьма озадачивающим явлением для всех, кто верит в какую-либо форму эволюции, особенно для тех, кто верит в исключительно постепенная эволюция» (17, с. 239).
239).
Болл пришел к выводу, что покрытосеменные впервые появились в высокогорных районах. Критически он утверждал (2, стр. 579), что, поскольку «только в крайне редких случаях растения из верхней горной области могут быть сохранены [окаменелыми]», долгая домеловая история покрытосеменных полностью отсутствует в летописи окаменелостей. пока цветущие растения позже не спустились на более низкие высоты (где окаменение было обычным явлением). Гипотеза Болла началась с понимания того, что уровни углекислого газа в атмосфере в каменноугольный период были чрезвычайно высокими (Болл утверждал, что в 20 раз выше, чем сейчас) и впоследствии снизились, в основном в результате образования угля (захоронения углерода).К этому Болл также предположил, что уровень углекислого газа снижается с высотой. Наконец, Болл предположил, что цветковые растения не могут выдерживать высокие уровни углекислого газа в атмосфере (в отличие от других групп растений). Таким образом, длительный период эволюции покрытосеменных растений (относящийся к каменноугольному периоду) должен был быть ограничен альпийским периодом, пока глобальные уровни углекислого газа в атмосфере не упали значительно (17).
Как заметил Хукер Дарвину в письме от 26 июля 1879 г. (происхождение: Библиотека Кембриджского университета, DAR 104: 128–130), «я думаю, что это очень неудовлетворительно во многих отношениях… и я уверен, что с вас уже достаточно Болл, которого мы обсудим при встрече.В печати Хукер был гораздо более дипломатичен (в опубликованных письменных комментариях Хукера после статьи 2): «Предположения [Болла] о происхождении рассматриваемых Флор, на которые повлияло присутствие этого газа в прежних условиях земной шар, у него [Хукера] перехватило дыхание». Дарвин тоже был настроен скептически, как он ясно дал понять Хукеру в своем отвратительном загадочном письме. «Безусловно, было бы большим шагом, если бы мы могли поверить, что высшие растения сначала могли жить только на высоком уровне; но пока экспериментально [доказано], что цикадовые, папоротники и т. д., могут выдерживать гораздо больше углекислоты, чем высшие растения, эта гипотеза кажется мне слишком опрометчивой». Тем не менее, на тему пропавшей летописи окаменелостей ранних покрытосеменных есть некоторые выдачи желаемого за действительное, потому что Дарвин отмечает, что «было бы большим шагом», если бы гипотеза Болла могла быть доказана.
Тем не менее, на тему пропавшей летописи окаменелостей ранних покрытосеменных есть некоторые выдачи желаемого за действительное, потому что Дарвин отмечает, что «было бы большим шагом», если бы гипотеза Болла могла быть доказана.
Гипотеза Болла об альпийском происхождении покрытосеменных заинтриговала Дарвина. В нем представлена та же самая аргументация, что и в его личных письмах: (1) внезапное появление разнообразных покрытосеменных растений в середине мелового периода было иллюзорным; (2) что длительный период эволюции покрытосеменных предшествовал тому, что тогда было известно с середины мелового периода; и (3) что не было ископаемых свидетельств постепенного разнообразия покрытосеменных растений до мелового периода.Единственная реальная разница между Боллом и Дарвином заключалась в том, что Болл предполагал, что длинная ранняя история покрытосеменных растений никогда не упоминалась в летописи окаменелостей, тогда как Дарвин постулировал, что они сохранились геологически на отдаленном острове или континенте, но массив суши исчез с лица земли. земли.
земли.
Альтернативное решение ужасной тайны
В отличие от мнения Дарвина о том, что первоначальная диверсификация покрытосеменных была постепенной, но не зафиксирована в палеонтологической летописи, альтернативное объяснение радиации цветковых растений среднего мелового периода было разработано и доведено до его сведения французским палеонтологом Гастоном де Сапорта (рис. .3). Дарвин открыл Сапорте (10 сентября 1876 г.; 7, стр. 93), что «относительно внезапного развития двудольных [ sic ] растений, которого также придерживается Хир, я, признаюсь, настроен скептически». В этом письме Дарвин повторяет свое обычное объяснение: «Конечно, это всего лишь догадка, но я полагаю, что эта большая группа растений должна была медленно развиваться в какой-то части земного шара, которая прежде была более изолированной от всех». другие регионы, чем какая-либо часть земли сейчас.Я всегда испытывал живой интерес к вашим наблюдениям за очень постепенными изменениями видов в течение позднего третичного периода, и я замечаю, что А. де Кандоль также был поражен этими наблюдениями, которые резко противоречат вере Геера в большие и резкие специфические изменения. изменения.»
де Кандоль также был поражен этими наблюдениями, которые резко противоречат вере Геера в большие и резкие специфические изменения. изменения.»
Сапорта, как и Дарвин, был озадачен кажущимся внезапным происхождением и быстрой диверсификацией покрытосеменных растений, что подтверждается летописью окаменелостей («un phénomène des plus curieux»).Однако, в отличие от Дарвина, Сапорта исходил из того, что быстрая скорость диверсификации покрытосеменных растений в середине мелового периода может быть реальной и может быть объяснена биологическими принципами. Важно отметить, что гипотеза Сапорты не требовала долгой незарегистрированной истории покрытосеменных растений до мелового периода.
Сапорта уже разработал общую теорию, которая могла обеспечить биологическую основу для быстрого разнообразия цветковых растений. Во втором разделе введения к Paléontologie Française, Plantes Jurassiques , 50 утверждал, что сильная взаимозависимость животных и растений требует понимания их различных связанных фаз эволюционной истории, и он описал храповой механизм совместной эволюции между животными и растениями. 7).В частности, Сапорта рассудил, что существенное отсутствие покрытосеменных растений в юрском периоде сделало невозможным эволюцию многих форм животных, особенно тех, которые были фитофагами. Как отмечают 7 и 13, 50 был первым, кто предположил критическую и взаимозависимую роль насекомых в появлении и разнообразии покрытосеменных растений.
7).В частности, Сапорта рассудил, что существенное отсутствие покрытосеменных растений в юрском периоде сделало невозможным эволюцию многих форм животных, особенно тех, которые были фитофагами. Как отмечают 7 и 13, 50 был первым, кто предположил критическую и взаимозависимую роль насекомых в появлении и разнообразии покрытосеменных растений.
В своей переписке с Чарльзом Дарвином Сапорта подробно остановился на теме коэволюционной взаимозависимости между насекомыми и цветковыми растениями — и на этот раз он связал свое прочтение летописи окаменелостей с вопросами, связанными с темпами диверсификации , самой сутью дарвиновской « гнусная тайна.В письме, известном своим блеском и проницательностью, Сапорта прямо предложил Дарвину (16 декабря 1877 г.; происхождение: Библиотека Кембриджского университета, DAR 177: 34; также обратите внимание, что транскрипция, использованная для этого перевода, отличается от опубликованной 7, стр. 98–99), что быстрая диверсификация покрытосеменных была, по сути, коэволюционной историей, связанной с происхождением многих основных групп насекомых: «Вы знаете, как замедленная эволюция двудольных всегда занимала меня как одну из самых любопытные явления, как из-за его огромной важности, так и из-за кажущейся скорости, с которой он раньше проявлялся. Теперь та роль, которую вы приписываете Насекомым в оплодотворении [опылении], вкупе с необходимостью скрещивания, объясняет все: прежнюю бедность растительного царства, сводившуюся так долго при отсутствии определенных категорий насекомых, к одним только анемофильным растениям , количество и разнообразие которых никогда не могло превысить определенного предела, и в которых питательные и сочные вещества никогда не были ни в изобилии, ни в достаточном разнообразии. Отсутствие сосущих насекомых во время юрского периода поразило меня так же, как и мистера Уайта.Heer. Я упомянул об этом отсутствии или его редкости в моем введении к юрской флоре, том. 1, стр. 53 и 54…
Теперь та роль, которую вы приписываете Насекомым в оплодотворении [опылении], вкупе с необходимостью скрещивания, объясняет все: прежнюю бедность растительного царства, сводившуюся так долго при отсутствии определенных категорий насекомых, к одним только анемофильным растениям , количество и разнообразие которых никогда не могло превысить определенного предела, и в которых питательные и сочные вещества никогда не были ни в изобилии, ни в достаточном разнообразии. Отсутствие сосущих насекомых во время юрского периода поразило меня так же, как и мистера Уайта.Heer. Я упомянул об этом отсутствии или его редкости в моем введении к юрской флоре, том. 1, стр. 53 и 54…
«Теперь можно очень хорошо понять, что покрытосеменные растения, у которых цветочные сочетания и скрещивания особей с особями и цветка с цветком зависят от роли насекомых, могли появиться и размножиться только под влиянием последних, а последних — для их часть могла стать многочисленной и активной [как опылители] и прилепиться к определенному определенному типу, по причине чего появление растений благоприятствовало их существованию; Таким образом, насекомые и растения были одновременно причиной и следствием благодаря их связи друг с другом, причем растения не могли разнообразиться без насекомых, а последние не могли давать много питателей пыльцой и нектаром до тех пор, пока царство растений оставалось бедным в устройстве и состоял почти исключительно из анемофильных растений 90–140 .
Дарвин немедленно ответил Сапорте (письмо от 24 декабря 1877 г.; 7, с. 109). «Ваша идея, что двудольные растения не развились в силу до тех пор, пока не развились сосущие насекомые, кажется мне великолепной. Я удивлен, что эта идея никогда не приходила мне в голову, но это всегда так, когда впервые слышишь новое и простое объяснение какого-то таинственного явления… [В]аша идея, которую, я надеюсь, ты опубликуешь, идет гораздо дальше и намного более важно…» Дарвин признал первостепенную важность гипотезы Сапорты и, таким образом, получил правдоподобный, действительно мощный механизм, объясняющий быстрые темпы ранней (имеющей в виду средний меловой период, по состоянию на 1877 г.) диверсификации покрытосеменных растений: ускоряющие эффекты совместной эволюции насекомых-опылителей и цветов, а также потенциальные преимущества усиленного ауткроссинга, связанного с энтомофилией.
Дарвин не забыл идеи Сапорты. Полтора года спустя в своем отвратительном таинственном письме к Джозефу Хукеру Дарвин сообщал: «Сапорта считает, что произошло удивительно быстрое развитие высших растений, как только [] насекомые, посещающие цветы, развились и благоприятствовали перекрестному скрещиванию». В 1881 г. (письмо к Джозефу Хукеру от 6 августа; 17, стр. 248) Дарвин снова вернулся к идеям Сапорты: «Поэтому меня очень заинтересовала точка зрения, которую Сапорта изложил мне несколько лет назад, очень подробно в MS, и который, я думаю, он с тех пор опубликовал, как я убедил его сделать, а именно.что, как только развились посещающие цветы насекомые, во второй половине вторичного периода был дан огромный толчок развитию высших растений благодаря внезапно образовавшемуся перекрестному опылению». Стоит отметить, что в этих двух письмах Дарвин сосредоточился на предполагаемых преимуществах ауткроссинга для содействия диверсификации, в то время как Сапорта в первую очередь подчеркивал коэволюционную динамику антофильных насекомых и цветковых растений. Сапорта подробно остановился на ранней совместной эволюции насекомых и покрытосеменных во втором томе L’Evolution du Règne Végétal (51).
В 1881 г. (письмо к Джозефу Хукеру от 6 августа; 17, стр. 248) Дарвин снова вернулся к идеям Сапорты: «Поэтому меня очень заинтересовала точка зрения, которую Сапорта изложил мне несколько лет назад, очень подробно в MS, и который, я думаю, он с тех пор опубликовал, как я убедил его сделать, а именно.что, как только развились посещающие цветы насекомые, во второй половине вторичного периода был дан огромный толчок развитию высших растений благодаря внезапно образовавшемуся перекрестному опылению». Стоит отметить, что в этих двух письмах Дарвин сосредоточился на предполагаемых преимуществах ауткроссинга для содействия диверсификации, в то время как Сапорта в первую очередь подчеркивал коэволюционную динамику антофильных насекомых и цветковых растений. Сапорта подробно остановился на ранней совместной эволюции насекомых и покрытосеменных во втором томе L’Evolution du Règne Végétal (51).
Таким образом, Дарвин, каким-то образом, который ранее не принимался во внимание, был, по крайней мере, частично ответственен за стимулирование публикации единственной наиболее принятой гипотезы относительно причины быстрого излучения цветковых растений, а именно их коэволюции с насекомыми (для продолжающихся дискуссий коэволюции покрытосеменных и насекомых см.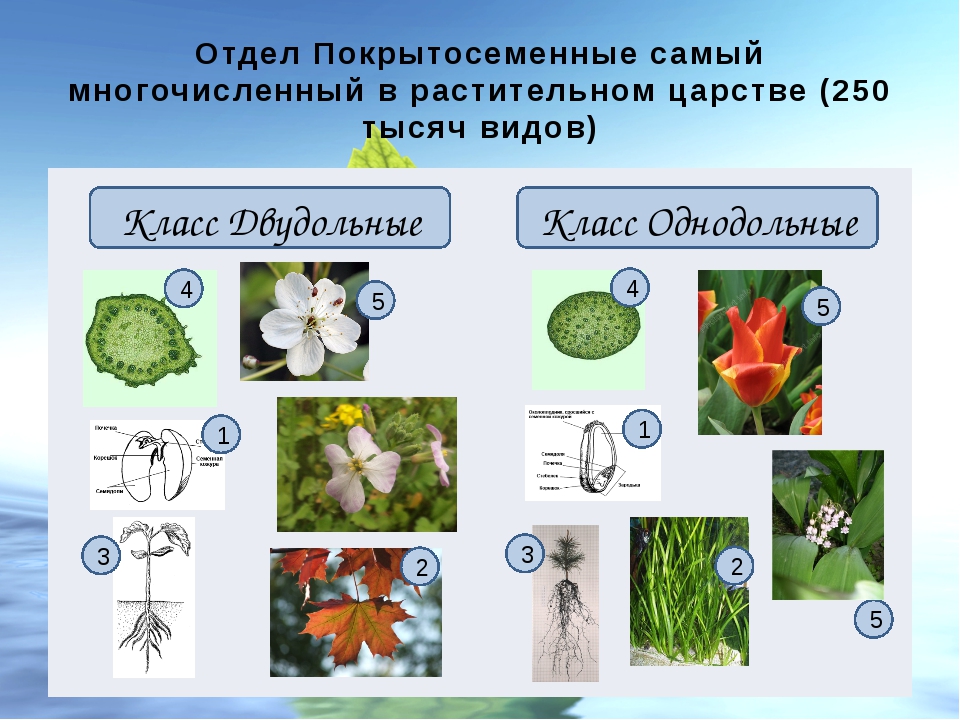 20; 46; 47; 10, 9; 43; 36; 49; 21; 31; 30; 52). Более того, решение Сапорты скептицизма Дарвина по поводу «внезапного развития» цветковых растений было единственным объяснением, которое Дарвин когда-либо использовал, чтобы потенциально объяснить быстрое разнообразие ранних покрытосеменных растений как реальность.
20; 46; 47; 10, 9; 43; 36; 49; 21; 31; 30; 52). Более того, решение Сапорты скептицизма Дарвина по поводу «внезапного развития» цветковых растений было единственным объяснением, которое Дарвин когда-либо использовал, чтобы потенциально объяснить быстрое разнообразие ранних покрытосеменных растений как реальность.
Включает ли в себя гнусная тайна происхождение покрытосеменных растений?
Ужасная тайна Дарвина обычно на протяжении 20-го и начала 21-го веков интерпретировалась как включающая в себя вопросы эволюции или трансформации признаков, связанных с происхождением (в отличие от их последующего разнообразия) цветковых растений от их не покрытосеменных предков. Как 58, с. 457) изложил дело (отражая точку зрения, восходящую к началу 1900-х годов): «Около ста лет назад Чарльз Дарвин называл происхождение покрытосеменных «отвратительной тайной».Современные исследования, хотя и пролили свет на многие проблемы, которые во времена Дарвина были загадочными и нерешенными, мало что сделали для прояснения этой проблемы. Она до сих пор занимает мысли многих ботаников, интересующихся эволюцией, к их недоумению и разочарованию. Причины этой трудности многообразны, но главным образом в несовершенстве летописи окаменелостей. Четкие доказательства происхождения и ранней эволюционной истории покрытосеменных должны состоять из ряда хорошо сохранившихся репродуктивных структур , соединяющих самые примитивные покрытосеменные растения с не покрытосеменными семенными растениями [курсив добавлен]…»
Она до сих пор занимает мысли многих ботаников, интересующихся эволюцией, к их недоумению и разочарованию. Причины этой трудности многообразны, но главным образом в несовершенстве летописи окаменелостей. Четкие доказательства происхождения и ранней эволюционной истории покрытосеменных должны состоять из ряда хорошо сохранившихся репродуктивных структур , соединяющих самые примитивные покрытосеменные растения с не покрытосеменными семенными растениями [курсив добавлен]…»
Хотя Дарвин был явно озадачен кажущимся внезапным происхождением и быстрой скоростью диверсификации покрытосеменных растений, он, по-видимому, не рассматривал в явном виде эволюцию многих уникальных характеристик цветковых растений (вопросы гомологии) от их не покрытосеменных предков.В основе его недоумения по поводу происхождения и ранней эволюции покрытосеменных определенно лежала проблема с летописью окаменелостей до покрытосеменных растений и филогении семенных растений. Обширная переписка Дарвина с палеоботаниками, описывающими растительную летопись юрского и мелового периодов, ясно показывает, что непосредственная предыстория цветковых растений интересовала его, по крайней мере косвенно.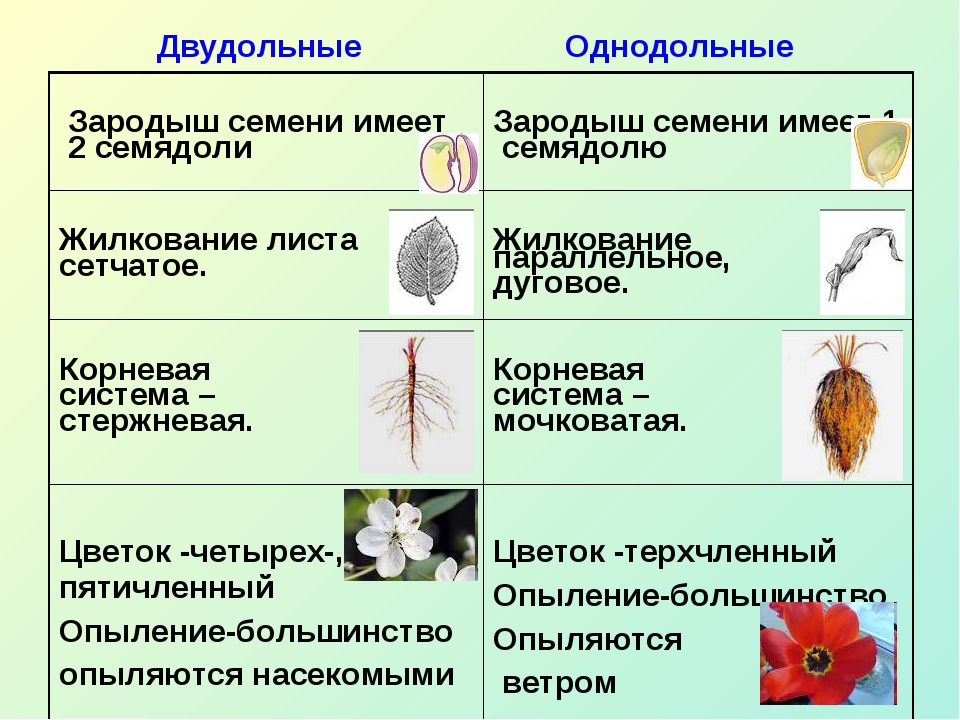 Тем не менее, Дарвин, по-видимому, не задумывался над преобразованиями специфических признаков, которые сыграли бы центральную роль в установлении определяющих признаков цветковых растений (например,g., цветок, плодолистик, зародышевый мешок и т. д.) — также нет свидетельств того, что он озаботился идентификацией предков цветковых растений. Принимая во внимание давнее признание Дарвином неадекватности летописи окаменелостей (обсуждается позже) и принятие им гипотезы о том, что цветковые растения могли развиваться в течение длительного периода времени в отдаленной области южного полушария, справедливо заключить, что что он просто не обратился к летописи окаменелостей за данными, которые могли бы говорить о гомологии и преобразованиях, которые привели к определяющим характеристикам покрытосеменных.
Тем не менее, Дарвин, по-видимому, не задумывался над преобразованиями специфических признаков, которые сыграли бы центральную роль в установлении определяющих признаков цветковых растений (например,g., цветок, плодолистик, зародышевый мешок и т. д.) — также нет свидетельств того, что он озаботился идентификацией предков цветковых растений. Принимая во внимание давнее признание Дарвином неадекватности летописи окаменелостей (обсуждается позже) и принятие им гипотезы о том, что цветковые растения могли развиваться в течение длительного периода времени в отдаленной области южного полушария, справедливо заключить, что что он просто не обратился к летописи окаменелостей за данными, которые могли бы говорить о гомологии и преобразованиях, которые привели к определяющим характеристикам покрытосеменных.
Эта отвратительная тайна была вызвана и в наше время в связи с филогенетическими отношениями между существующими семенными растениями и, в частности, с идентификацией ближайших ныне живущих родственников покрытосеменных растений. Хотя анализ филогении семенных растений в последние годы Дарвина шел полным ходом (например, Эрнст Геккель, писавший в 1870-х и 1880-х годах в своей основополагающей работе «История сотворения: или развитие Земли и ее обитателей» , рассматривал гнеталес как наиболее тесно связанный с покрытосеменными), в его трудах и переписке эта тема не затрагивается.
Хотя анализ филогении семенных растений в последние годы Дарвина шел полным ходом (например, Эрнст Геккель, писавший в 1870-х и 1880-х годах в своей основополагающей работе «История сотворения: или развитие Земли и ее обитателей» , рассматривал гнеталес как наиболее тесно связанный с покрытосеменными), в его трудах и переписке эта тема не затрагивается.
Дарвин был по существу агностиком в вопросе взаимоотношений внутри семенных растений и признал это в конце 1861 г. в письме Джозефу Хукеру (17, с. 281; письмо от 28 декабря 1861 г.): «Я небрежно писал о ценности фанерогам; я думал о том, что подгруппы, казалось, смешивались друг с другом гораздо больше, чем с большинством классов животных». В 1862 и 1863 гг., когда Джозеф Хукер занимался интенсивным изучением вегетативных, репродуктивных и эмбриологических особенностей Welwitschia (33), Дарвин обратил внимание (письмо от 6 октября 1862 г.; 4) на мнение Хукера о том, что Gnetales (и в особенности Welwitschia ) можно рассматривать как промежуточные по своему признаку между голосеменными и покрытосеменными («действительно кажется грандиозным случаем соединить две такие группы»).
Имеющиеся данные убедительно свидетельствуют о том, что Дарвин не уделял должного внимания вопросу о предполагаемых родственниках покрытосеменных растений и их потенциальном эволюционном значении. В то время как происхождение покрытосеменных растений от не покрытосеменных семенных растений остается проблематичным даже по сей день (например, 24; 41; 5), вопросы гомологии и трансформации признаков, вопросы идентификации предков и ближайших родственников покрытосеменных не были предметом обсуждения. центральная часть любопытства и разочарования Дарвина в отношении эволюционной истории цветковых растений.
Natura non facit saltum, суть отвратительной тайны Дарвина
В течение прошлого века дарвиновская «отвратительная тайна» стала в широком смысле синонимом сложностей и часто кажущихся неразрешимыми вопросов, связанных с предысторией, происхождением и ранними фазами эволюционной истории покрытосеменных. Тем не менее, немногие из этих тем, если они вообще есть, поскольку они относятся конкретно к цветковым растениям, по-видимому, занимали главное место в уме Дарвина в любой момент его жизни.И это приводит к последнему и важному набору соображений: действительно ли ужасная тайна Дарвина касается покрытосеменных растений?
Тем не менее, немногие из этих тем, если они вообще есть, поскольку они относятся конкретно к цветковым растениям, по-видимому, занимали главное место в уме Дарвина в любой момент его жизни.И это приводит к последнему и важному набору соображений: действительно ли ужасная тайна Дарвина касается покрытосеменных растений?
В заключительной главе каждого издания «Происхождение видов » (с заменой всего одного-двух слов) Дарвин писал: «Поскольку естественный отбор действует исключительно путем накопления незначительных, последовательных, благоприятных вариаций, он не может производить больших или внезапных изменений». модификация; он может действовать только очень короткими и медленными шагами. Отсюда канон «Natura non facit saltum [природа не делает скачка]», который каждое новое дополнение к нашим знаниям стремится сделать более строго правильным…» (14, первое издание, с.471; по существу аналогично в 15, шестое издание, стр. 413–414). Однако появляющаяся летопись окаменелостей покрытосеменных растений в 1860-х и 1870-х годах представила Дарвину наиболее явные доказательства, противоречащие его градуалистской точке зрения на макроэволюционные изменения.
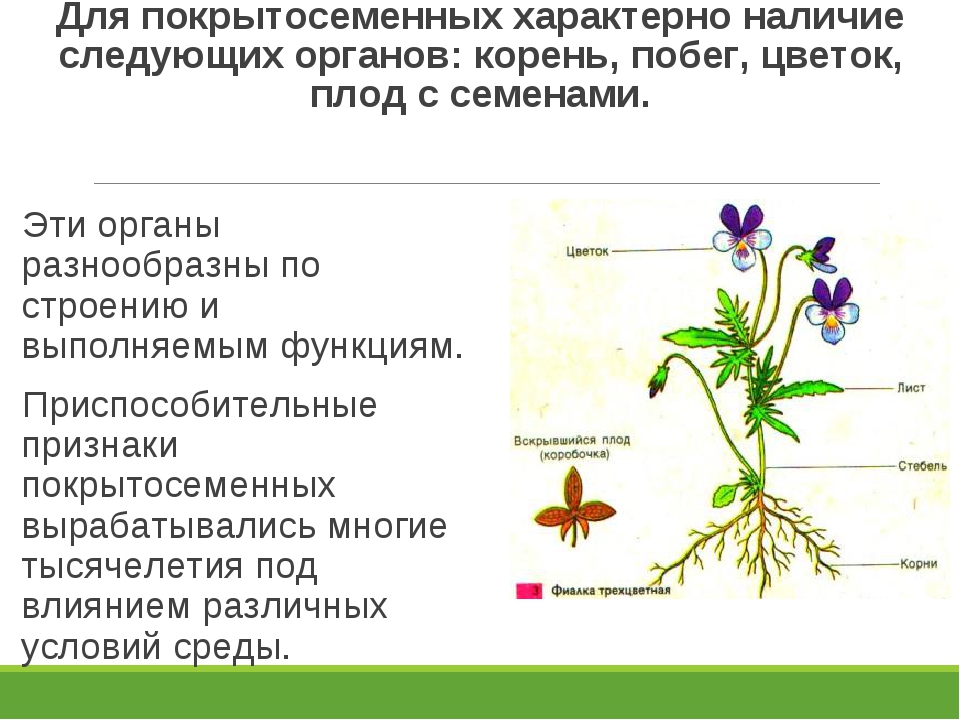 «Неожиданный характер, с которым целые группы видов внезапно появляются в определенных формациях, указывался некоторыми палеонтологами, например, Агассисом, Пикте, и никем более настойчиво, чем профессором Седжвиком, как фатальное возражение против веры в существование трансмутация видов.Если бы многочисленные виды, принадлежащие к одним и тем же родам или семействам, действительно возникли одновременно, то этот факт был бы фатальным для теории происхождения с медленными изменениями в результате естественного отбора. Ибо развитие группы форм, все из которых произошли от какого-то одного прародителя, должно было быть чрезвычайно медленным процессом; и прародители должны были жить задолго до их видоизмененных потомков» (14, первое издание, стр. 302; по существу то же самое в 15, шестое издание, стр. 282).
«Неожиданный характер, с которым целые группы видов внезапно появляются в определенных формациях, указывался некоторыми палеонтологами, например, Агассисом, Пикте, и никем более настойчиво, чем профессором Седжвиком, как фатальное возражение против веры в существование трансмутация видов.Если бы многочисленные виды, принадлежащие к одним и тем же родам или семействам, действительно возникли одновременно, то этот факт был бы фатальным для теории происхождения с медленными изменениями в результате естественного отбора. Ибо развитие группы форм, все из которых произошли от какого-то одного прародителя, должно было быть чрезвычайно медленным процессом; и прародители должны были жить задолго до их видоизмененных потомков» (14, первое издание, стр. 302; по существу то же самое в 15, шестое издание, стр. 282).
Искусным тактическим маневром Дарвин подготовил почву для решения этого предполагаемого «фатального» вызова его теории эволюции по происхождению с постепенными модификациями. «[Мы] постоянно переоцениваем совершенство геологической летописи и делаем ложный вывод, поскольку определенные роды или семейства не были обнаружены ниже определенной стадии, что они не существовали до этой стадии. Мы постоянно забываем, насколько велик мир по сравнению с областью, на которой наши геологические формации были тщательно исследованы; мы забываем, что группы видов могли давно существовать где-то еще и медленно размножаться, прежде чем они вторглись на древние архипелаги Европы и Соединенных Штатов» (14, первое издание, с.302; по существу аналогично в 15, шестое издание, с. 282). Дарвин продолжает: «Теперь я приведу несколько примеров, чтобы проиллюстрировать предыдущие замечания и показать, насколько мы склонны ошибаться, предполагая, что целые группы видов возникли внезапно» (14, первое издание, стр. 302; идентично в 15, шестое издание, стр. 283). Этим Дарвин предпринимает эмпирическую контратаку против капризов летописи окаменелостей, описывая то, что рассматривалось (до публикации «Происхождение видов ») как кажущееся внезапным происхождение различных представителей млекопитающих, птиц, сидячих усоногих, костистых ракообразных.
«[Мы] постоянно переоцениваем совершенство геологической летописи и делаем ложный вывод, поскольку определенные роды или семейства не были обнаружены ниже определенной стадии, что они не существовали до этой стадии. Мы постоянно забываем, насколько велик мир по сравнению с областью, на которой наши геологические формации были тщательно исследованы; мы забываем, что группы видов могли давно существовать где-то еще и медленно размножаться, прежде чем они вторглись на древние архипелаги Европы и Соединенных Штатов» (14, первое издание, с.302; по существу аналогично в 15, шестое издание, с. 282). Дарвин продолжает: «Теперь я приведу несколько примеров, чтобы проиллюстрировать предыдущие замечания и показать, насколько мы склонны ошибаться, предполагая, что целые группы видов возникли внезапно» (14, первое издание, стр. 302; идентично в 15, шестое издание, стр. 283). Этим Дарвин предпринимает эмпирическую контратаку против капризов летописи окаменелостей, описывая то, что рассматривалось (до публикации «Происхождение видов ») как кажущееся внезапным происхождение различных представителей млекопитающих, птиц, сидячих усоногих, костистых ракообразных. рыб и трилобитов.В каждом случае он утверждал, что то, что всего несколько лет назад казалось внезапным происхождением различных членов каждой группы, теперь может быть показано как имеющее значительный предшествующий отчет о переходных окаменелостях, совместимый с его градуалистскими взглядами на трансмутацию.
рыб и трилобитов.В каждом случае он утверждал, что то, что всего несколько лет назад казалось внезапным происхождением различных членов каждой группы, теперь может быть показано как имеющее значительный предшествующий отчет о переходных окаменелостях, совместимый с его градуалистскими взглядами на трансмутацию.
Эти отрывки из книги «Происхождение видов» предвосхищают стратегию, которую позже применил Дарвин, чтобы раскрыть отвратительную тайну кажущегося внезапным и географически широко распространенного появления разнообразных покрытосеменных растений в летописи окаменелостей.Идеи Дарвина в книге «Происхождение видов » явно основаны на предположении, что биологические инновации и разнообразие происходят постепенно, как он позже утверждал в случае с цветковыми растениями. Более того, Дарвин отмечает, что ранняя эволюция линии может быть географически ограничена (удалена от Европы и Северной Америки, где в то время проводилась большая часть палеонтологии) и, как таковая, не обнаружена в летописи окаменелостей. Параллель с затерянным островом или континентом в южном полушарии в случае покрытосеменных растений точна.
Параллель с затерянным островом или континентом в южном полушарии в случае покрытосеменных растений точна.
К 1870-м годам Дарвин пережил серию интеллектуальных и тактических битв за различные группы многоклеточных животных, которые, казалось бы, внезапно появились в летописи окаменелостей. Критически важно, что его склонность в пользу градуалистских объяснений трансмутации возникла в период, отмеченный растущей, но крайне неполной летописью окаменелостей большинства групп организмов. Это привело его к поиску объяснений кажущегося внезапным происхождения и разнообразия основных родословных животных и растений, представленных в известных летописях окаменелостей того времени.Тем не менее, в каждом случае, 90 139 и только в одном 90 140, геологические и эволюционные «проблемы» Дарвина решались по мере того, как палеонтологическим исследованиям удавалось заполнить недостающие пробелы. У Дарвина были все основания полагать, что время на его стороне.
Когда Дарвин начал критически исследовать (и переписываться) летопись окаменелостей покрытосеменных, от него ускользнули доказательства постепенной диверсификации цветковых растений до их резкого и разнообразного появления (как это было известно в 1870-х годах) в середине мелового периода. Еще в 1878 году (16 февраля) Сапорта повторил Дарвину, ссылаясь на летопись окаменелостей цветковых растений (происхождение: Библиотека Кембриджского университета, DAR 177: 35): «Поскольку нет никаких следов двудольных до середины мелового периода… первые поддающиеся определению типы с этого времени кажутся украшенными своими отличительными характеристиками и фиксированными в своих основных чертах. Это отсутствие первоначальных [переходных] форм делает переход к основной [группе] — матерям, из которых должен был возникнуть весь класс… проблема тем более неясная. это не то же самое для млекопитающих, где можно наблюдать почти каждый переход и последовательность, ведущую от одного типа к другой дефинитивной группе.Дарвин пришел к своей отвратительной тайне не из-за особого или программного любопытства к широкомасштабной эволюционной истории цветковых растений, а скорее потому, что покрытосеменные среди всех групп живых организмов представляли собой величайший постоянный вызов его взглядам на темпы развития.
Еще в 1878 году (16 февраля) Сапорта повторил Дарвину, ссылаясь на летопись окаменелостей цветковых растений (происхождение: Библиотека Кембриджского университета, DAR 177: 35): «Поскольку нет никаких следов двудольных до середины мелового периода… первые поддающиеся определению типы с этого времени кажутся украшенными своими отличительными характеристиками и фиксированными в своих основных чертах. Это отсутствие первоначальных [переходных] форм делает переход к основной [группе] — матерям, из которых должен был возникнуть весь класс… проблема тем более неясная. это не то же самое для млекопитающих, где можно наблюдать почти каждый переход и последовательность, ведущую от одного типа к другой дефинитивной группе.Дарвин пришел к своей отвратительной тайне не из-за особого или программного любопытства к широкомасштабной эволюционной истории цветковых растений, а скорее потому, что покрытосеменные среди всех групп живых организмов представляли собой величайший постоянный вызов его взглядам на темпы развития. эволюционных инноваций и его ясное признание того, что летопись окаменелостей остается существенно неполной.
эволюционных инноваций и его ясное признание того, что летопись окаменелостей остается существенно неполной.
Всю свою жизнь Дарвин был одержим адаптацией и кумулятивными небольшими изменениями, которые создают новые инновации в биологии.Вопросы эволюционной истории (например, гомология и трансформация признаков) отдельных групп организмов, хотя, несомненно, и интересовали Дарвина в целом, редко, если вообще когда-либо, были мотивирующими элементами его исследований. Его письма ясно показывают, что он не особенно интересовался филогении покрытосеменных или семенных растений, а также не был одержим отслеживанием ископаемой предыстории цветковых растений. Нет никаких указаний на то, что Дарвин интересовался гомологией цветка и его органов (как, например, Гёте в 1790 г.).
Отвратительная тайна Чарльза Дарвина никогда не касалась цветковых растений как таковых. Отвратительная загадка Чарльза Дарвина была, по сути, одной из или : как объяснить периоды кажущейся резкой и очень ускоренной диверсификации, связанные с происхождением основных групп организмов — в данном случае и лишь случайно покрытосеменных.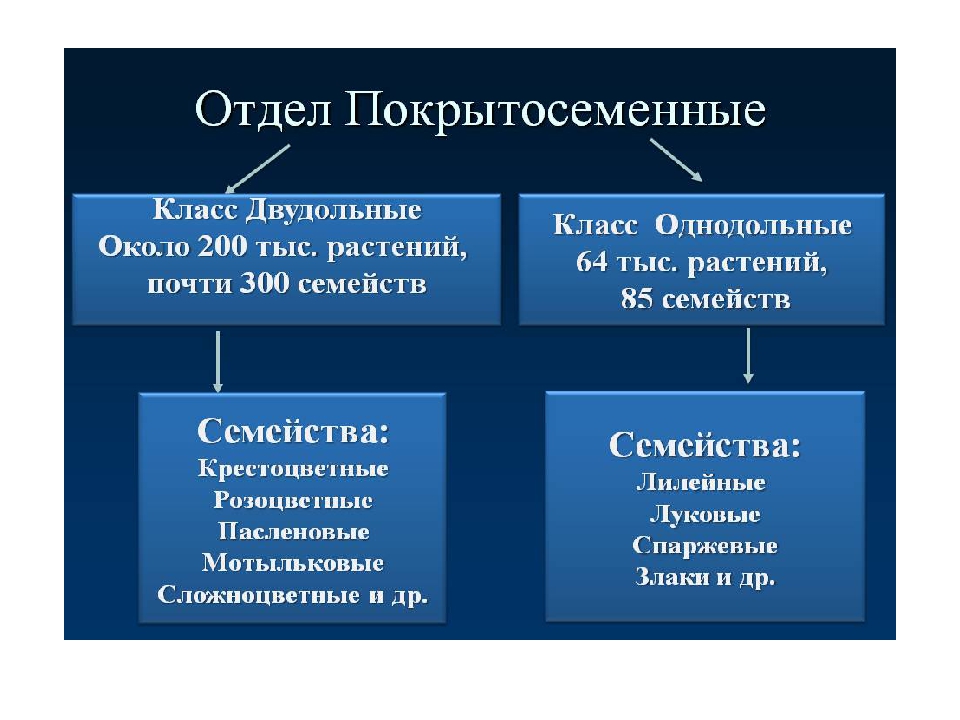 Именно поэтому Дарвин возвращался к теме сальтационистских взглядов Хира в своей переписке как с самим Хиром, так и с Сапортой.Дарвин не мог согласиться с тем, что темпы эволюционных инноваций могут быть такими быстрыми, и покрытосеменные растения (известные в 1870-х и 1880-х годах), по-видимому, представляли значительное исключение из этого глубоко укоренившегося представления.
Именно поэтому Дарвин возвращался к теме сальтационистских взглядов Хира в своей переписке как с самим Хиром, так и с Сапортой.Дарвин не мог согласиться с тем, что темпы эволюционных инноваций могут быть такими быстрыми, и покрытосеменные растения (известные в 1870-х и 1880-х годах), по-видимому, представляли значительное исключение из этого глубоко укоренившегося представления.
Заключительные замечания
Когда Дарвин отправился в свою могилу (на самом деле, не в могилу, где он рассчитывал быть погребенным на кладбище Даун вместе со своей семьей, а скорее в Вестминстерское аббатство), палеонтологическая летопись все еще представляла собой свидетельство кажущегося внезапным происхождения и чрезвычайно быстрой эволюционной и биогеографической диверсификации цветущие растения.Таким образом, стоит кратко остановиться на том, была ли «отвратительная тайна» Дарвина , особенно в отношении покрытосеменных растений, «разрешена» в промежутке между 1882 годом и настоящим временем.
Теперь мы знаем, что самые ранние фазы диверсификации покрытосеменных, о чем свидетельствует летопись окаменелостей, демонстрируют период относительно постепенной и упорядоченной диверсификации до середины мелового периода (см. обзор 25). Эта раннемеловая летопись покрытосеменных, в флоре которой преобладали мелкоцветковые виды, была совершенно неизвестна палеонтологам времен Дарвина (или даже 30 лет назад).Таким образом, самое раннее проявление покрытосеменных, как теперь понимается, кажется значительно менее резким, чем в палеонтологической летописи Хеера и Сапорты, описанной в 1870-х и 1880-х годах. Во многих отношениях недавно обнаруженная запись о ранних покрытосеменных согласуется с моделями, которые Дарвин находил утешением, когда он воспринял открытия ранних переходных окаменелостей млекопитающих, птиц, сидячих ракушек, костистых рыб и трилобитов в книге «Происхождение видов» . Очевидно, такие предположения открыты для обсуждения, но Дарвин вполне мог бы удовлетвориться добавлением покрытосеменных к своему списку примеров, «чтобы показать, насколько мы склонны ошибаться, предполагая, что целые группы видов возникли внезапно», если бы он имел доступ к наша текущая запись цветковых растений раннего мела.
Примечательно, что и Дарвин, и Болл были правы, заключая, что ранняя летопись окаменелостей покрытосеменных в том виде, в каком она существовала в 1870-х годах, была принципиально неполной. Оба прекрасно понимали, что кажущееся внезапным происхождение многих дифференцированных (производных) членов большой группы организмов без летописи окаменелостей переходных предков не согласуется с эволюционной схемой происхождения видов и разнообразием более высокого уровня. По сути, оба они предсказали, что до середины мелового периода должен был существовать период диверсификации покрытосеменных растений.Их предсказания в отношении покрытосеменных полностью подтвердились.
Несмотря на капризы стратиграфии середины XIX века, Дарвину была очевидна сущность излучения различных клад покрытосеменных от середины до позднего мела (и раннего третичного периода). Текущие качественные показатели по-прежнему указывают на высокие темпы морфологического, экологического и таксономического разнообразия покрытосеменных в период с середины позднего мелового и раннего третичного периодов (44; 8; 61; 37; 9; 11, 3; 38; 40; 25). .Было высказано предположение, что значительное расширение летописи окаменелостей покрытосеменных от среднего до позднего мелового и раннего третичного периодов в течение прошлого века могло усугубить «проблему» высоких темпов диверсификации, с которой столкнулся Дарвин (13). Однако стоит иметь в виду, что в 1860-х и 1870-х годах преобладала точка зрения, выдвинутая лордом Кельвином (Уильямом Томсоном), что возраст Земли составляет от десятков до сотен миллионов лет (35). Учитывая, что в настоящее время известно, что только меловой период имеет продолжительность порядка 80 миллионов лет, неясно, рассматривал ли Дарвин диверсификацию покрытосеменных от среднего до позднего мела (как теперь понимается) как особенно неприятную для своего градуалиста. взгляды.В любом случае, это вопрос, принципиально отличный от вопроса о «по-видимому, очень внезапном или резком развитии высших растений», с которым столкнулся Дарвин и который мотивировал его переписку с Хукером, Хиром и Сапортой.
.Было высказано предположение, что значительное расширение летописи окаменелостей покрытосеменных от среднего до позднего мелового и раннего третичного периодов в течение прошлого века могло усугубить «проблему» высоких темпов диверсификации, с которой столкнулся Дарвин (13). Однако стоит иметь в виду, что в 1860-х и 1870-х годах преобладала точка зрения, выдвинутая лордом Кельвином (Уильямом Томсоном), что возраст Земли составляет от десятков до сотен миллионов лет (35). Учитывая, что в настоящее время известно, что только меловой период имеет продолжительность порядка 80 миллионов лет, неясно, рассматривал ли Дарвин диверсификацию покрытосеменных от среднего до позднего мела (как теперь понимается) как особенно неприятную для своего градуалиста. взгляды.В любом случае, это вопрос, принципиально отличный от вопроса о «по-видимому, очень внезапном или резком развитии высших растений», с которым столкнулся Дарвин и который мотивировал его переписку с Хукером, Хиром и Сапортой.
Таким образом, кажущееся внезапным происхождение различных групп многоклеточных животных, а также покрытосеменных растений, было предметом горячих споров между креационистами и эволюционистами 19-го века. Дарвин хорошо понимал интеллектуальную дилемму, связанную с появлением различных представителей различных групп организмов без переходной ископаемой предыстории.Тем не менее было бы ошибкой делать вывод из вырезанных Дарвином слов «отвратительная тайна», сделанных по отношению к происхождению и быстрому разнообразию цветковых растений, что он специально и программно интересовался историей эволюции покрытосеменных растений. Отвратительная тайна Дарвина в конечном счете и, как показывают его переписка и публикации, не связана с покрытосеменными растениями как таковыми. Отвратительная загадка Дарвина заключается в его отвращении к тому, что эволюция может быть одновременно быстрой и потенциально даже скачкообразной; речь идет о его опасениях, что кажущиеся внезапными появления хорошо дифференцированных членов группы организмов могут быть использованы для поддержки креационистской программы; и это действительно отражение ограничений, с которыми столкнулись Дарвин и другие эволюционисты его эпохи, пытаясь прочесть еще очень неполную летопись окаменелостей жизни на Земле.
Дарвин хорошо понимал интеллектуальную дилемму, связанную с появлением различных представителей различных групп организмов без переходной ископаемой предыстории.Тем не менее было бы ошибкой делать вывод из вырезанных Дарвином слов «отвратительная тайна», сделанных по отношению к происхождению и быстрому разнообразию цветковых растений, что он специально и программно интересовался историей эволюции покрытосеменных растений. Отвратительная тайна Дарвина в конечном счете и, как показывают его переписка и публикации, не связана с покрытосеменными растениями как таковыми. Отвратительная загадка Дарвина заключается в его отвращении к тому, что эволюция может быть одновременно быстрой и потенциально даже скачкообразной; речь идет о его опасениях, что кажущиеся внезапными появления хорошо дифференцированных членов группы организмов могут быть использованы для поддержки креационистской программы; и это действительно отражение ограничений, с которыми столкнулись Дарвин и другие эволюционисты его эпохи, пытаясь прочесть еще очень неполную летопись окаменелостей жизни на Земле.
Примитивные цветковые растения | Управление по управлению влажными тропиками
В конце юрского периода растения прошли еще одну веху эволюции с появлением цветковых растений в летописи окаменелостей. Это были предшественники современных растений с их более совершенными механизмами опыления. Начиная с Западной Гондваны, на территории современной Западной Африки и восточной части Южной Америки, цветковые растения (называемые покрытосеменными) распространились по суперконтиненту как раз в тот момент, когда он начал распадаться в начале мелового периода.На каком-то этапе считалось, что покрытосеменные могли возникнуть в северном Квинсленде из-за большого количества примитивных покрытосеменных, которые мы имеем сегодня, но летопись окаменелостей показала обратное.
Древние родословные
Филогенетическая переоценка мировой флоры (с использованием молекулярного секвенирования) позволила классифицировать 28 почти базальных (или «примитивных») линий цветковых растений. Влажные тропики могут претендовать на 16 из этих почти базальных семейств, все из которых находятся в местах обитания тропических лесов.Эти 16 семейств содержат 46 родов и 175 видов. (см. таблицу ниже). Тропические леса влажных тропиков поддерживают большое количество семейств, которые можно описать как происходящие от линий, разветвляющихся у основания генеалогического древа покрытосеменных. Эти семейства, роды и виды заключают в себе в генетическом смысле большую часть эволюционной истории. Это свидетельствует о долгой истории постоянно сохраняющихся тропических лесов во влажных тропиках, что стало возможным благодаря климатическим условиям и наличию прохладных влажных горных убежищ, где многие растения пережили более засушливые времена.
Влажные тропики могут претендовать на 16 из этих почти базальных семейств, все из которых находятся в местах обитания тропических лесов.Эти 16 семейств содержат 46 родов и 175 видов. (см. таблицу ниже). Тропические леса влажных тропиков поддерживают большое количество семейств, которые можно описать как происходящие от линий, разветвляющихся у основания генеалогического древа покрытосеменных. Эти семейства, роды и виды заключают в себе в генетическом смысле большую часть эволюционной истории. Это свидетельствует о долгой истории постоянно сохраняющихся тропических лесов во влажных тропиках, что стало возможным благодаря климатическим условиям и наличию прохладных влажных горных убежищ, где многие растения пережили более засушливые времена.
Вы можете прочитать подробности о древних линиях цветковых растений в «Переоценке влажных тропиков Квинсленда на основе примитивных растений».
Несколько примитивных растений
Чтобы пробудить ваш интерес, вот лишь несколько примитивных цветковых растений. Смотрите также сюжет о Зеленом Динозавре.
Смотрите также сюжет о Зеленом Динозавре.
Австробайлея
Примитивная лиана, австробайлея, редкая и эндемичная для влажных тропиков (нигде больше не встречается).Это единственный вид в своем семействе, обитающий в очень влажных тропических лесах от низких до возвышенностей. Austrobaileya scandens — лиана с пологом, высота которой составляет около 15 метров. Его листья необычного голубовато-зеленого цвета, но его цветы выдают, что это один из самых ранних представителей цветковых растений. Его цветочные части расположены по спирали, а лепестки бледно-зеленого цвета. Мухи — единственные опылители, и чтобы их привлечь, цветы пахнут гниющей рыбой! Плоды абрикосового цвета продолговатой формы размером до 70 мм на 50 мм свисают с лозы и содержат скопление семян в форме каштана.
Родная гуава или медный лавр
Многие растения тропических лесов очень нуждаются в защите от самоопыления, и, поскольку особи могут быть рассеяны друг от друга с большим количеством густой листвы между ними, некоторые виды развили сложные системы, гарантирующие, что они не могут самоопыляться. . Местная гуава или медный лавр ( Eupomatia laurina ) имеет именно такую систему. Его опылителем является жук (что объясняет, почему его цветок имеет слегка ужасный запах), но его цветок распускается в два этапа.На каждом дереве женские части цветка обнажаются первыми в начале дня. Затем эта часть цветка закрывается, а мужские части (тычинки) вытягиваются. Это гарантирует, что ни одно дерево не может самоопыляться. Аборигенная гуава представляет собой реликтовый, примитивный вид, все семейство которого состоит только из одного рода, состоящего из трех видов.
. Местная гуава или медный лавр ( Eupomatia laurina ) имеет именно такую систему. Его опылителем является жук (что объясняет, почему его цветок имеет слегка ужасный запах), но его цветок распускается в два этапа.На каждом дереве женские части цветка обнажаются первыми в начале дня. Затем эта часть цветка закрывается, а мужские части (тычинки) вытягиваются. Это гарантирует, что ни одно дерево не может самоопыляться. Аборигенная гуава представляет собой реликтовый, примитивный вид, все семейство которого состоит только из одного рода, состоящего из трех видов.
Перечное дерево
Перечное дерево ( Tasmannia мембрана ) получило свое название из-за вкуса его семян, а не его мягких плодов.Это дерево принадлежит к одному из самых ранних известных семейств цветковых растений — Winteraceae, что делает его настоящим реликтом тропического леса. Перечное дерево больше похоже на большой кустарник, чем на дерево, достигающее только трех метров в высоту. Он имеет общую характеристику с менее развитыми саговниками — у него отдельные мужские и женские цветки на разных растениях.
Он имеет общую характеристику с менее развитыми саговниками — у него отдельные мужские и женские цветки на разных растениях.
Миристина
Ducula bicolor , научное название голубя Торресова пролива или имперского пестрого голубя, означает «пожиратель Myristica» или, чаще говоря, «пожиратель мускатного ореха».Myristica — это род, содержащий только три вида во влажных тропиках, один из которых мы знаем как мускатный орех Квинсленда. Плоды яйцевидной формы цвета ржавчины размером 20 мм являются любимой пищей императорского голубя-пестрого, отсюда и его научное название. Квинслендский мускатный орех ( Myristica globulosa subsp. muelleri ) может использоваться для коммерческого производства пряного мускатного ореха, но вместо него используется аналогичный вид с Соломоновых островов. ( Myristica fragrans с Молуккских островов дает коммерческие специи мускатного ореха (семена) и мускатного ореха (ариллы).Это 15-метровое дерево является еще одним примером примитивного цветкового растения, а также имеет отдельные деревья для мужских и женских цветков.
Эволюционный анализ семейства генов дегидринов покрытосеменных растений выявил три группы ортологов, связанных со специфическими белковыми доменами
Беспристрастная полногеномная идентификация дегидринов в геномах Viridiplantae , YnKn-SKn- и Kn-DHN), мы провели полногеномный поиск для идентификации репертуаров генов DHN в 56 геномах видов Viridiplantae, включая представителей хлорофитов и стрептофитов.Первоначальный скрининг был проведен с использованием профиля скрытой марковской модели (HMM), определенного для белков семейства дегидринов (Pfam00257)
31 . Неожиданно, когда мы проанализировали извлеченные последовательности, мы заметили, что хорошо известные KS-DHN, такие как дегидрин HIRD11 из Arabidopsis thaliana (At1g54410) 32 и ZmDHN13 из Zea mays 8 не были обнаружены алгоритм. Это побудило нас предположить, что Pfam00257 недостаточно чувствителен для распознавания KS-DHN.Чтобы преодолеть это ограничение, мы построили три разных профиля HMM: один (KS-HMM) с использованием последовательностей KS-DHN из геномов покрытосеменных, идентифицированных поиском Blastp с использованием A. thaliana HIRD11, и два других профиля (F-HMM и Y-HMM). на основе белков покрытосеменных, принадлежащих к группам ортологичных F- и Y-DHN, недавно описанных 17 .
thaliana HIRD11, и два других профиля (F-HMM и Y-HMM). на основе белков покрытосеменных, принадлежащих к группам ортологичных F- и Y-DHN, недавно описанных 17 . После поиска с помощью Pfam00257 и групповых профилей HMM мы восстановили в общей сложности 305 неповторяющихся последовательностей из геномов репрезентативных видов мохообразных (4), ликофитов (1), голосеменных (3) и покрытосеменных (36) ( Дополнительные данные S1).Никаких последовательностей не было получено из геномов водорослей, что позволяет предположить, что DHN могли появиться в наземных растениях 7 .
Примечательно, что профиль KS-HMM показал повышенную чувствительность при распознавании гомологов DHN, поскольку он смог идентифицировать 92,2% белков DHN, в то время как HMM на основе Pfam00257 идентифицировал 83% (рис. 1). Из 62 DHN, использующих архитектуру KS, только 17 удалось получить с использованием профиля Pfam00257, что подтверждает его низкую эффективность при распознавании KS-DHN. В то время как F-HMM имеет лучшую производительность и может распознать 24 KS-DHN, только профиль KS-HMM смог получить все KS-DHN. Действительно, 32 KS-DHN можно было получить только с использованием профиля KS-HMM. И наоборот, профиль KS-HMM не смог распознать 24 последовательности DHN, которые были идентифицированы другими профилями HMM. Никакие DHN не были получены исключительно с помощью Pfam00257, что указывает на то, что групповые профили HMM необходимы для тщательного поиска белков DHN в геномах.
В то время как F-HMM имеет лучшую производительность и может распознать 24 KS-DHN, только профиль KS-HMM смог получить все KS-DHN. Действительно, 32 KS-DHN можно было получить только с использованием профиля KS-HMM. И наоборот, профиль KS-HMM не смог распознать 24 последовательности DHN, которые были идентифицированы другими профилями HMM. Никакие DHN не были получены исключительно с помощью Pfam00257, что указывает на то, что групповые профили HMM необходимы для тщательного поиска белков DHN в геномах.
DHN, идентифицированные в геномах Viridiplantae с помощью различных профилей HMM.Последовательности, извлеченные из геномов видов с использованием профилей F-HMM, Y-HMM, KS-HMM, построенных в этом исследовании, и профиля Pfam00257 для белков семейства дегидринов, отображаются в виде диаграммы Венна. Белые числа указывают количество KS-DHN, присутствующих в каждом подмножестве. Обратите внимание, что большинство KS-DHN не восстанавливаются с помощью Pfam00257.
Анализ консервативного белкового мотива и классификация базы данных дегидринов
Мы использовали цМем для проверки наличия известных дегидриновых мотивов (K-Y-F- и S-сегменты) и для обнаружения предполагаемых новых мотивов (дополнительный рис.С1). Консервативные мотивы и количество обнаруженных DHN в структурных подгруппах показаны на рис. 2.
Рисунок 2 Идентификация консервативных белковых мотивов и структурная классификация DHN. ( a ) Представление LOGO различных консервативных мотивов, обнаруженных цМемом в наборе DHN объективной базы данных. ( b ) Количество членов каждой структурной подгруппы покрытосеменных DHN, идентифицированных в объективной базе данных. Мы выделили структурные подгруппы FSK2 и FSK3 в соответствии со Strimbeck 6 .Все классифицированные DHN, принадлежащие к 19 семействам покрытосеменных, перечислены в дополнительных данных S1. Пунктирный узор указывает на однодольные растения, а закрашенный — на двудольные.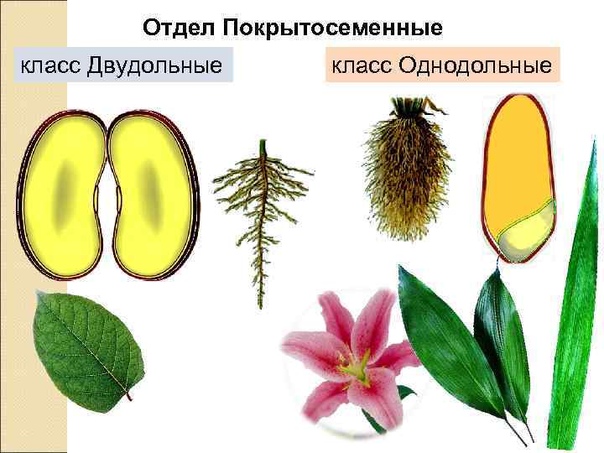
Мы подтвердили наличие K-сегмента в 302 из 307 дегидринов, идентифицированных поиском гомологии на основе профилей HMM. MEME не смог распознать K-сегмент в нескольких белках, все из которых относятся к видам, не относящимся к покрытосеменным. Однако все эти белки обладают вырожденным, менее консервативным K-сегментом, а также другими мотивами DHN, что указывает на то, что они являются настоящими DHN.Так обстоит дело, например, с DHN из плауновидного S. moellendorffii и голосеменного Ginkgo biloba (рис. 3).
Рисунок 3 Консервация KS-DHN в сосудистых растениях. Множественное выравнивание последовательностей KS-DHN (HKS-DHN) репрезентативных покрытосеменных растений ( A. trichopoda , S. bicolor , N. nucifera , A. thaliana , M. truncatula truncatula ) (плауновидный S. moellendorffii и голосеменный G.biloba ), выполненный с помощью T-Coffee и визуализированный с помощью Jalview. Обозначены H-, K-, B и S-сегменты. Отметим, что общая структура белков сохраняется во всех группах сосудистых растений.
Обозначены H-, K-, B и S-сегменты. Отметим, что общая структура белков сохраняется во всех группах сосудистых растений.
Мы идентифицировали 75 DHN, несущих уникальный F-сегмент, расположенный в N-концевой области белков, которые мы отнесли к структурной подгруппе FSKn-DHN. Сегмент F, предсказанный с помощью нашей базы данных, аналогичен сегменту, описанному Стримбеком 6 (рис. 2а). Важно отметить, что даже поиск с конкретным профилем F-HMM не смог идентифицировать FSKn-DHN в геномах четырех мохообразных и одного плауновидного, но мы нашли их в трех голосеменных, включенных в это исследование, Picea abies , Picea glauca. и Г.biloba , подтверждая, что эта группа возникла в семенных растениях. Мы наблюдали расширение группы FSKn-DHN в кладе Pinaceae в соответствии с предыдущими наблюдениями 33 , но только три DNH были идентифицированы у G. biloba , два с F-сегментом (FSK2 и FK2) и третий — KS-DHN (см. Ниже). У покрытосеменных подгруппа FSKn-DHN в основном состоит из белков FSK2 и FSK3 (рис. 2b), но, что интересно, только FSK3-DHN обнаружены у однодольных и у ранней дивергентной эвдикоты Nelumbo nucifera , а также у базальных покрытосеменные А.trichopoda и N. colorata (дополнительные рисунки S2 – S3), что позволяет предположить, что FSK2-DHN могли возникнуть из предковых FSK3-DHN.
2b), но, что интересно, только FSK3-DHN обнаружены у однодольных и у ранней дивергентной эвдикоты Nelumbo nucifera , а также у базальных покрытосеменные А.trichopoda и N. colorata (дополнительные рисунки S2 – S3), что позволяет предположить, что FSK2-DHN могли возникнуть из предковых FSK3-DHN.
Мы обнаружили в общей сложности 101 DHN, содержащих от одной до трех копий Y-сегмента на последовательность в N-концевом положении, все они у покрытосеменных растений. Большинство белков принадлежат к подгруппе YnSKn, тогда как последовательности без S-сегмента (YnKn) составляют лишь 15%. Как у однодольных, так и у двудольных мы обнаружили YSKn и Y2SKn-DHN, в то время как Y3SKn- и YnKn-DHN, по-видимому, встречаются только у двудольных (рис.2б; Дополнительные рис. С4–С6). Поразительно, что Phi-сегмент, обнаруженный в нашей базе данных, соответствует N-концевой области ранее определенного GT-сегмента 7 и является высококонсервативным у однодольных YSKn.
Мы идентифицировали в общей сложности 62 KS-DHN в геномах растений, все из которых имеют новый N-концевой мотив (H-сегмент, см. ниже). Интересно, что профиль KS-HMM позволил нам идентифицировать KS-DHN в геноме плауновидного S. mollendorfii и голосеменного G.biloba , но в геномах хвойных P. abies и P. glauca KS-DHN не идентифицированы. Белки представляют собой типичное расположение мотивов KS с K-сегментом, за которым следует участок, богатый лизином (B-сегмент), и S-сегмент, характерный для этой структурной подгруппы (S2-сегмент) (рис. 3). Это первый случай, когда KS-DHN идентифицированы у не покрытосеменных видов, что указывает на то, что эта группа DHN возникла на ранней стадии эволюции наземных растений.
У покрытосеменных все проанализированные виды обладали одним или двумя генами KS-DHN, за исключением Glycine max с четырьмя генами и двух видов Malpighiales, Salix purpurea и Populus trichocarpa с шестью и тремя KS- DHN соответственно. Белки Malpigiales уникальны, поскольку они содержат несколько повторов K-сегмента, перемежающихся последовательностями, богатыми глицином (Phi-сегмент), а S2-сегмент отсутствует (дополнительная рис. S8).
Белки Malpigiales уникальны, поскольку они содержат несколько повторов K-сегмента, перемежающихся последовательностями, богатыми глицином (Phi-сегмент), а S2-сегмент отсутствует (дополнительная рис. S8).
Что касается структурных подгрупп Kn- и SKn-DHN, то их представленность в сосудистых растениях была незначительной и, в конечном счете, они филогенетически родственны другим структурным подгруппам DHN (см. ниже). Напротив, большинство из восемнадцати несосудистых DHN, которые мы идентифицировали в геномах мхов P.patens (шесть белков), Ceratodon purpureus (шесть) , Sphagnum fallax (три) и печеночница Marchantia polymorpha (три) относятся к Kn-структурной подгруппе. Исключением является атипичный DHN, содержащий серию повторяющихся мотивов, напоминающих Y-сегмент из P. patens (PpDHNA) 34 и C. purpureus (дополнительный рисунок S10). Филогенетический анализ указывает на наличие пяти групп ортологов DHN у P.patens и C.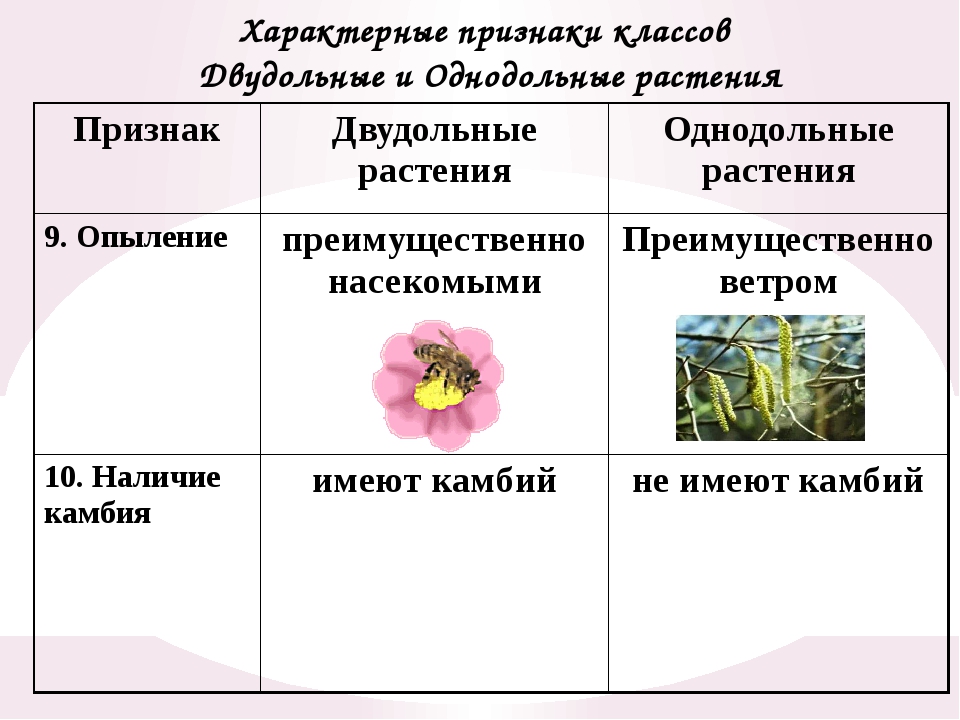 purpureus (дополнительные рисунки S9–S10), что отражает филогенетическую близость клад Funariidae и Dicranidae 35 , в то время как DHN из S. fallax (Sphagnophytina) не группировались с другими мхи. DHN печеночника M. polymorpha не обнаруживают очевидной гомологии с DHN мха за пределами K-сегмента.
purpureus (дополнительные рисунки S9–S10), что отражает филогенетическую близость клад Funariidae и Dicranidae 35 , в то время как DHN из S. fallax (Sphagnophytina) не группировались с другими мхи. DHN печеночника M. polymorpha не обнаруживают очевидной гомологии с DHN мха за пределами K-сегмента.
H-сегмент представляет собой новый консервативный мотив, присутствующий во всех KS-дегидринах
Наш анализ цМемов выявил высококонсервативный мотив, ранее не описанный, в N-концевой области всех проанализированных KS-DHN покрытосеменных.Этот сегмент из 15 остатков характеризуется комбинацией гидрофобных аминокислот Ile и Leu с амфипатическими аминокислотами Lys и Glu, обрамленными двумя Gly в положениях 3 и 15, консервативными в 91% и 87% KS-DHN соответственно (рис. 2). ). Кроме того, наблюдается высокая консервативность Lys (97%) и Ile (97%) в центральных положениях 7 и 8. Остатки Ile в положениях 4 и 5 менее консервативны и часто заменяются другими гидрофобными аминокислотами, такими как Phe, Вал или Мет. Два остатка His обнаружены в положениях 6 и 13 в 56% и 77% KS-DHN соответственно.KS-DHN характеризуются последовательностями, обогащенными аминокислотами His, что отражено в названии HIRD11 для A. thaliana KS-DHN, что означает Hi стидин- R ich D омаин 11 кДа 36 . Поскольку этот новый мотив, по-видимому, является сигнатурой KS-DHN, мы предлагаем назвать его Н-сегментом , , что отражает особую природу белков этого типа.
Два остатка His обнаружены в положениях 6 и 13 в 56% и 77% KS-DHN соответственно.KS-DHN характеризуются последовательностями, обогащенными аминокислотами His, что отражено в названии HIRD11 для A. thaliana KS-DHN, что означает Hi стидин- R ich D омаин 11 кДа 36 . Поскольку этот новый мотив, по-видимому, является сигнатурой KS-DHN, мы предлагаем назвать его Н-сегментом , , что отражает особую природу белков этого типа.
Внутренне неупорядоченные белки (IDP), такие как дегидрины, не имеют единой четко определенной третичной структуры в нативных условиях.С помощью программного обеспечения IUPred2A мы идентифицировали участки неупорядоченного белка в репрезентативных KS-DHN покрытосеменных растений (H-DHN) (рис. 4). Интересно, что области, охватывающие H- и K-сегменты, соответствуют двум уникальным областям, которые демонстрируют пониженную тенденцию к беспорядку.
Рисунок 4 Графики прогнозирования беспорядка для репрезентативных H-DHN покрытосеменных растений (KS-DHN). С помощью программы IUPred2A было предсказано нарушение белков A. trichopoda , S. bicolor и A. thaliana H-DHN.Обратите внимание, что все белки прогнозируются как полностью неупорядоченные (оценка > 0,5), однако области, соответствующие H- и K-мотивам, демонстрируют уменьшенную тенденцию к беспорядку. Обозначены H-, K-, B и S-сегменты.
С помощью программы IUPred2A было предсказано нарушение белков A. trichopoda , S. bicolor и A. thaliana H-DHN.Обратите внимание, что все белки прогнозируются как полностью неупорядоченные (оценка > 0,5), однако области, соответствующие H- и K-мотивам, демонстрируют уменьшенную тенденцию к беспорядку. Обозначены H-, K-, B и S-сегменты.
K- и S-сегменты KS-DHN обладают некоторыми особыми характеристиками по сравнению с FSK и YSK-DHN (рис. 2a и дополнительные рисунки S2–S7). Позиция 15 занята Ile почти во всех KS-DHN вместо Lys, который обычно присутствует в других DHN. Несмотря на то, что аминокислоты Ile и Leu обычно считаются консервативными, есть свидетельства того, что эти аминокислоты не всегда взаимозаменяемы, что влияет на сродство и специфичность белок-белковых и белок-мембранных взаимодействий 37 , что потенциально может привести к функциональной диверсификации KS-DHN, модулируя поведение K-сегмента.В отличие от других DHN, KS-дегидрины не показывают явного преобладания Pro в положении 16; вместо этого His является наиболее часто встречающейся аминокислотой в этом положении. Несмотря на эти различия, предсказанная структура α-спирали, ограниченная консервативными аминокислотами Gly K-сегмента, сохраняется в KS-DHN (рис. 4).
Несмотря на эти различия, предсказанная структура α-спирали, ограниченная консервативными аминокислотами Gly K-сегмента, сохраняется в KS-DHN (рис. 4).
Что касается S-сегментов, которые характеризуются участком остатков Ser, существуют различия в длине участка Ser-аминокислоты и соседних аминокислот между KS-DHN и другими DHN.Ядро из 6-9 остатков Ser обычно заканчивается отрицательно заряженными аминокислотами Asp или Glu во всех структурных подгруппах DHN. С другой стороны, триада Leu-His-Arg, которая предшествует отрезку Ser, которая высоко консервативна во всех FSK-DHN и в большинстве YSK-DHN, не обнаруживается в KS-DHN. На рисунке 2a показан консенсус S-сегмента для FSK и YSK-DHN (сегмент S1), а также консенсус, обнаруженный в KS-DHN (сегмент S2, см. Также выравнивание на дополнительных рисунках S2 и S7). Было показано, что S-сегмент всех типов DHN является горячей точкой для фосфорилирования киназами 8,38,39 , а различия между S1- и S2-сегментами могут приводить к различной специфичности киназ. Например, триада Leu-His-Arg составляет часть последовательности узнавания для киназ SnRK2 40, которые, как было недавно продемонстрировано, фосфорилируют дегидринов A. thaliana ERD4 и ERD10 в ответ на осмотический стресс 41.
Например, триада Leu-His-Arg составляет часть последовательности узнавания для киназ SnRK2 40, которые, как было недавно продемонстрировано, фосфорилируют дегидринов A. thaliana ERD4 и ERD10 в ответ на осмотический стресс 41.
Мотив, богатый Lys из 11 остатков, постоянно обнаруживался во всех KS-DHN, а также во всех FSK2 и большинстве FSK3-DHN (дополнительные рисунки S1 – S3; рис. 5). В KS- и FSK-DHN Lys-богатый мотив расположен между S-сегментом и K-сегментом, в то время как в том же положении YK- и YSK-DHN обычно имеют последовательность RRKK или RRKKK, обрамленную остатками Gly, мотив, напоминающий одночастные сигналы ядерной локализации 42,43 .В заключение, KS-DHN можно лучше описать как имеющие структурную организацию H-K-S, где H является недавно описанным сегментом, присутствующим исключительно в этой группе DHN.
Рисунок 5 Некорневое филогенетическое древо белков DHN покрытосеменных растений. Аминокислотные последовательности белков DHN из покрытосеменных были выровнены, и с помощью IQ-TREE была создана реконструкция филогенетических взаимоотношений приблизительно с максимальной вероятностью. Ветки с поддержкой начальной загрузки более 90% отмечены фиолетовой точкой.Мотивы каждого белка обозначены цветными прямоугольниками, как указано. Обратите внимание, что все H-DHN (KS-DHN) сгруппированы вместе в ветви с высокой поддержкой, и большинство F- и Y-DHN также сгруппированы вместе, образуя три группы. Белки DHN из покрытосеменных A. trichopoda (красный) и эвдикот N. nucifera (синий) показаны, чтобы показать, что они обладают одним DHN в каждой группе. Дерево не имеет корней и показывает относительную связанность последовательностей, что означает, что положение кажущегося корня произвольно.
Аминокислотные последовательности белков DHN из покрытосеменных были выровнены, и с помощью IQ-TREE была создана реконструкция филогенетических взаимоотношений приблизительно с максимальной вероятностью. Ветки с поддержкой начальной загрузки более 90% отмечены фиолетовой точкой.Мотивы каждого белка обозначены цветными прямоугольниками, как указано. Обратите внимание, что все H-DHN (KS-DHN) сгруппированы вместе в ветви с высокой поддержкой, и большинство F- и Y-DHN также сгруппированы вместе, образуя три группы. Белки DHN из покрытосеменных A. trichopoda (красный) и эвдикот N. nucifera (синий) показаны, чтобы показать, что они обладают одним DHN в каждой группе. Дерево не имеет корней и показывает относительную связанность последовательностей, что означает, что положение кажущегося корня произвольно.
Филогенетический анализ выявил три основные группы DHN у покрытосеменных
Идентифицировав мотивы, характеризующие KS-структурную подгруппу, мы попытались сделать вывод о филогенетических взаимоотношениях между идентифицированными белками DHN. Мы решили использовать только DHN покрытосеменных для построения филогенетического дерева из-за редкой таксономической выборки DHN других наземных растений, а также из-за присущей трудности выравнивания последовательностей DHN более отдаленных видов, которые имеют более расходящиеся доменные структуры.Мы использовали принцип максимального правдоподобия, реализованный в IQ-TREE 23 , и оценили статистическую устойчивость с помощью метода начальной загрузки.
Мы решили использовать только DHN покрытосеменных для построения филогенетического дерева из-за редкой таксономической выборки DHN других наземных растений, а также из-за присущей трудности выравнивания последовательностей DHN более отдаленных видов, которые имеют более расходящиеся доменные структуры.Мы использовали принцип максимального правдоподобия, реализованный в IQ-TREE 23 , и оценили статистическую устойчивость с помощью метода начальной загрузки.
Результирующее дерево можно условно разделить на три ветви или группы (рис. 5). Все KS-DHN, характеризующиеся наличием H-сегмента (H-DHN), группируются в ветвь с высокой поддержкой бутстрапа (100%). Остальные DHN разделены на две ветви, одна из которых содержит большинство DHN, содержащих F-сегмент (FSKn), а другая содержит DHN, несущие Y-сегмент (YnSKn, YnKn).Интересно, что несколько DHN, которые содержат только K-сегмент или комбинацию K- и S-сегментов, расположены внутри ветвей F или Y, что указывает на то, что эти DHN фактически принадлежат к любой из этих групп. В целом филогенетическое дерево предполагает, что каждый DHN покрытосеменных принадлежит к одному из трех филогенетических семейств, в основном отличающихся наличием H-, F- или Y-сегментов. Этот вывод подкрепляется наблюдением DH растений в ключевых филогенетических позициях. Так, Amborella trichopoda , относящаяся к сестринской группе по отношению ко всем другим покрытосеменным («базальным» покрытосеменным растениям), обладает тремя DHN, каждая из которых принадлежит к группам H, F или Y (рис.5). Точно так же три DHN из Nelumbo nucifera , которые принадлежат к сестринской группе всех эвдикотов, также каждая из которых помещена в группы H, F и Y. В целом, филогенетические результаты предполагают, что эти три группы DHN присутствовали с самого начала эволюции покрытосеменных.
В целом филогенетическое дерево предполагает, что каждый DHN покрытосеменных принадлежит к одному из трех филогенетических семейств, в основном отличающихся наличием H-, F- или Y-сегментов. Этот вывод подкрепляется наблюдением DH растений в ключевых филогенетических позициях. Так, Amborella trichopoda , относящаяся к сестринской группе по отношению ко всем другим покрытосеменным («базальным» покрытосеменным растениям), обладает тремя DHN, каждая из которых принадлежит к группам H, F или Y (рис.5). Точно так же три DHN из Nelumbo nucifera , которые принадлежат к сестринской группе всех эвдикотов, также каждая из которых помещена в группы H, F и Y. В целом, филогенетические результаты предполагают, что эти три группы DHN присутствовали с самого начала эволюции покрытосеменных.
H-DHN принадлежат к отдельному синтеническому сообществу покрытосеменных
Несмотря на то, что описанное выше филогенетическое дерево разделяет DHN покрытосеменных растений на три группы, большое количество различных мотивов и их дивергентное расположение в DHN затрудняет выравнивание последовательностей и снижает достоверность филогенетической реконструкции. Поскольку анализ синтении генов-ортологов может дать важные подсказки об эволюции геномов и семейств генов 44 , мы провели анализ геномного соседства (микросинтении) генов DHN, чтобы подкрепить наши филогенетические результаты.
Поскольку анализ синтении генов-ортологов может дать важные подсказки об эволюции геномов и семейств генов 44 , мы провели анализ геномного соседства (микросинтении) генов DHN, чтобы подкрепить наши филогенетические результаты.
Недавно Артур и др. 17 проанализировали геномы генов DHN в геномах растений и выявили два основных синтенных блока (или сообщества) среди покрытосеменных, соответствующие DHN, содержащим мотивы F (сообщество 1) и Y (сообщество 2).Их анализ, однако, не включал DHN группы KS, предположительно из-за сложности извлечения этих последовательностей с использованием мотива Pfam PF00257. Чтобы убедиться, что DHN, содержащие H-сегмент, также являются частью синтетического сообщества, мы сравнили 40 генов, окружающих уникальный H-DHN базального покрытосеменного, A. trichopoda , с геномными окрестностями локусов H-DHN водяной лилии. Nymphaea colorata 45 , эвдикот базальный священный лотос, с.ш.NUCIFERA 46 , Begume , Medicago Truncatula 47 47 , модельный завод A. Thaliana (Hird11) 48 и монокот трава сорго Биколор 49 . Все эти виды обладают только одним паралогом H-DHN, за исключением M. truncatula , у которого их два (дополнительные данные S1). Мы выбрали A. trichopoda в качестве основы для сравнения синтении, поскольку это цветковое растение принадлежит к родственной линии всех других покрытосеменных (Amborellales), не подвергалось дупликации всего генома, которая затронула другие линии, и его геном демонстрирует консервативную синтению с другими покрытосеменными растениями. 50,51 .Как показано на рис. 6, 17 генов, которые окружают H-DHN A. trichopoda (LOC18421535), также присутствуют вокруг гена H-DHN N. colorata , принадлежащего к группе Nymphaeales, которая является сестринской линией для всех покрытосеменных, кроме Amborellales 52 . Меньшее количество консервативных генов присутствует вокруг генов H-DHN эвдикотов N. nucifera , A. thaliana ( HIRD11 ) и M.
Thaliana (Hird11) 48 и монокот трава сорго Биколор 49 . Все эти виды обладают только одним паралогом H-DHN, за исключением M. truncatula , у которого их два (дополнительные данные S1). Мы выбрали A. trichopoda в качестве основы для сравнения синтении, поскольку это цветковое растение принадлежит к родственной линии всех других покрытосеменных (Amborellales), не подвергалось дупликации всего генома, которая затронула другие линии, и его геном демонстрирует консервативную синтению с другими покрытосеменными растениями. 50,51 .Как показано на рис. 6, 17 генов, которые окружают H-DHN A. trichopoda (LOC18421535), также присутствуют вокруг гена H-DHN N. colorata , принадлежащего к группе Nymphaeales, которая является сестринской линией для всех покрытосеменных, кроме Amborellales 52 . Меньшее количество консервативных генов присутствует вокруг генов H-DHN эвдикотов N. nucifera , A. thaliana ( HIRD11 ) и M. truncatula и однодольных S.биколор (рис. 6). Микросинтения генов H-DHN других покрытосеменных растений также консервативна (не показано). H-DHN обладают двумя экзонами, при этом вся кодирующая область содержится в первом экзоне, а второй экзон не кодирует, в то время как F- и Y-DHN обычно имеют два кодирующих экзона (не показано). Консервативность экзон-интронной структуры также указывает на общее происхождение H-DHN. В заключение, микросинтения генов H-DHN сохраняется у покрытосеменных растений, что указывает на их истинный ортологический статус и общее эволюционное происхождение.
truncatula и однодольных S.биколор (рис. 6). Микросинтения генов H-DHN других покрытосеменных растений также консервативна (не показано). H-DHN обладают двумя экзонами, при этом вся кодирующая область содержится в первом экзоне, а второй экзон не кодирует, в то время как F- и Y-DHN обычно имеют два кодирующих экзона (не показано). Консервативность экзон-интронной структуры также указывает на общее происхождение H-DHN. В заключение, микросинтения генов H-DHN сохраняется у покрытосеменных растений, что указывает на их истинный ортологический статус и общее эволюционное происхождение.
Анализ микросинтении генов H-DHN покрытосеменных растений. Геномное соседство гена H-DHN A. trichopoda (LOC18421535) сравнивается с соседством других покрытосеменных растений. Гены H-DHN обозначены черными точками, а цветовой код указывает на гомологичные гены, присутствующие у других видов. Серые точки обозначают гены, присутствующие только у A. trichopoda . Некоторые промежуточные гены у видов, отличных от A. trichopoda , не показаны для ясности.
trichopoda . Некоторые промежуточные гены у видов, отличных от A. trichopoda , не показаны для ясности.
Наряду с одним геном H-DHN геном A. trichopoda содержит два других гена DHN (LOC18424350 и LOC18426770). Как упоминалось выше, в нашем филогенетическом дереве LOC18424350 сгруппирован вместе с F-DHN, а LOC18424350 с Y-DHN (рис. 5). Любопытно, что мотивы F и Y этих белков сильно вырождены и не сразу распознаются программой цМем. Сравнение геномных соседей LOC18424350 и LOC18426770 A. trichopoda с F- и Y-DHN N.colorata и N. nucifera , которые также имеют только три гена DHN, демонстрирует консервацию микросинтении вокруг этих генов (дополнительные данные S2), подтверждая, что LOC18424350 и LOC18426770 принадлежат синтетическим сообществам F- и Y-DHN соответственно.
Таким образом, очевидно, что гены DHN покрытосеменных можно в целом разделить на три синтетических сообщества, каждое из которых характеризуется, среди прочего, наличием мотива H, F или Y. Мы предлагаем называть эти ортологичные группы F-дегидринами (сообщество 1), Y-дегидринами (сообщество 2) и H-дегидринами (сообщество 3).Присутствие только трех генов дегидринов в геномах растений, принадлежащих к группам, которые дивергировались на ранней стадии эволюции покрытосеменных, как Amborella и Nymphaea , или на ранней стадии эволюции эвдикотов, как N. nucifera , предполагает, что геномы Первые цветковые растения имели по одному гену H-, F- и Y-дегидрина. Последующие дупликации всего генома эвдикотов и однодольных значительно увеличили репертуар этих генов, особенно тех, которые кодируют F- и Y-дегидрины.
Мы предлагаем называть эти ортологичные группы F-дегидринами (сообщество 1), Y-дегидринами (сообщество 2) и H-дегидринами (сообщество 3).Присутствие только трех генов дегидринов в геномах растений, принадлежащих к группам, которые дивергировались на ранней стадии эволюции покрытосеменных, как Amborella и Nymphaea , или на ранней стадии эволюции эвдикотов, как N. nucifera , предполагает, что геномы Первые цветковые растения имели по одному гену H-, F- и Y-дегидрина. Последующие дупликации всего генома эвдикотов и однодольных значительно увеличили репертуар этих генов, особенно тех, которые кодируют F- и Y-дегидрины.
Каждая группа ортологичных DHN обладает отличительными биохимическими свойствами гидрофилина
Мы сравнили биохимические и биофизические свойства DHN покрытосеменных из групп H-, F- и Y-ортологичных, а именно такие характеристики, как молекулярная масса (MW), изоэлектрическая точка (pI ), а также параметры, связанные с гидрофилиновым характером белков (дополнительные данные S1). Каждая ортологическая группа DHN имеет различное распределение MW с характерной статистической медианой (рис.7а). H-DHN являются наименьшими DHN с самым узким диапазоном молекулярной массы (10–16 кДа), что свидетельствует о том, что количество остатков и доменная структура в этой группе относительно постоянны. F-DHN также имеют компактное распределение молекулярной массы (18–35 кДа), в то время как Y-DHN представляют собой основную подгруппу белков с молекулярной массой 10–25 кДа и ряд DHN более 30 кДа, принадлежащих однодольным растениям. Высокая молекулярная масса этой последней подгруппы связана не с увеличением количества консервативных доменов Y или K, а с наличием длинных областей, богатых глием (дополнительные рис.С4–С6). Большинство H-DHN имеют кислые значения pI, при этом у некоторых видов обнаружены нейтральные и основные изоформы (рис. 7b). F-DHN имеют очень однородный кислотный профиль pI с одномодальным распределением между 5 и 6. Напротив, Y-DHN демонстрируют бимодальное распределение, состоящее из двух основных подгрупп DHN с основными и кислотными значениями pI, и меньшей подгруппы с pI ценности, близкие к нейтральности.
Каждая ортологическая группа DHN имеет различное распределение MW с характерной статистической медианой (рис.7а). H-DHN являются наименьшими DHN с самым узким диапазоном молекулярной массы (10–16 кДа), что свидетельствует о том, что количество остатков и доменная структура в этой группе относительно постоянны. F-DHN также имеют компактное распределение молекулярной массы (18–35 кДа), в то время как Y-DHN представляют собой основную подгруппу белков с молекулярной массой 10–25 кДа и ряд DHN более 30 кДа, принадлежащих однодольным растениям. Высокая молекулярная масса этой последней подгруппы связана не с увеличением количества консервативных доменов Y или K, а с наличием длинных областей, богатых глием (дополнительные рис.С4–С6). Большинство H-DHN имеют кислые значения pI, при этом у некоторых видов обнаружены нейтральные и основные изоформы (рис. 7b). F-DHN имеют очень однородный кислотный профиль pI с одномодальным распределением между 5 и 6. Напротив, Y-DHN демонстрируют бимодальное распределение, состоящее из двух основных подгрупп DHN с основными и кислотными значениями pI, и меньшей подгруппы с pI ценности, близкие к нейтральности.
Распределение биохимических и биофизических свойств ДГН покрытосеменных.Графики рассеяния показывают распределение молекулярной массы ( a ), изоэлектрической точки ( b и f ), содержания глицина ( c ), индекса GRAVY ( d ), индекса складки ( e ) или содержание глицина, индекс GRAVY и Fold одновременно ( г ) в ортологичных или структурных подгруппах DHN. Члены трех ортологических групп DHN имеют цветовую кодировку: Y- (зеленый), F- (оранжевый) и H-DHN (фиолетовый).
Что касается содержания глицина, как F-, так и H-DHN представляют собой компактное и однородное распределение процентного содержания остатков глицина (рис.7в). DHN с самым низким процентным содержанием глицина (около 10%) являются F-DHN. Примечательно, что DHN структурной подгруппы FSK3 характеризуются наличием множества участков пролина, которые могут играть роль, эквивалентную роли глицина, с точки зрения нарушения структуры белка (дополнительные рисунки S2 – S3).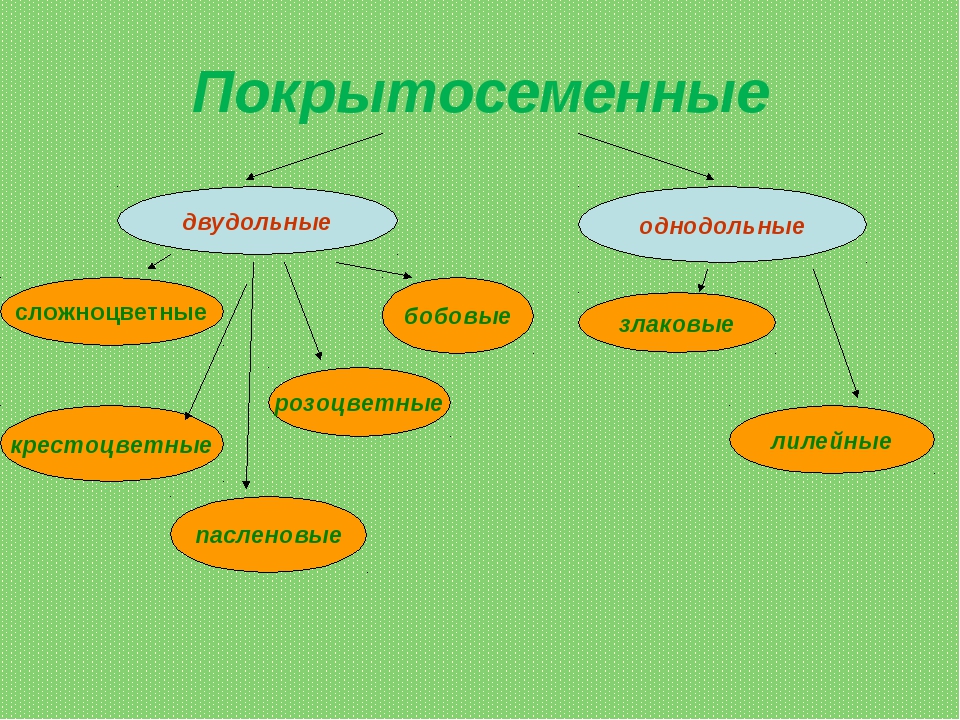 Примечательно, что больший разброс процентного содержания остатков глицина наблюдается в Y-DHN (5–35%).
Примечательно, что больший разброс процентного содержания остатков глицина наблюдается в Y-DHN (5–35%).
Все DHN имеют отрицательный индекс GRAVY, что указывает на характерную гидрофильность этих белков (рис.7г). Y-DHN включают наименее гидрофильные белки, тогда как H-DHN являются наиболее гидрофильными, с индексами GRAVY в диапазоне от -2,8 до -1,3. Атипичные H-DHN из S. purpurea и P. trichocarpa (Malpighiales) являются наименее гидрофильными в этой группе с индексом около -1,3. Баллы для F-DHN имеют промежуточный диапазон, перекрывающийся с другими группами. Мы также оценили Fold Index DHN, используя алгоритм FoldIndex 53 . Распределение индекса складки DHN аналогично распределению индекса GRAVY, при этом H-DHN являются наиболее развернутыми, в то время как F- и Y-DHN имеют менее развернутый характер (рис.7д).
При анализе распределения pI в пяти традиционных структурных подгруппах ДГН можно отметить, что бимодальный характер, наблюдаемый в ДГН SK- и К-типа, сильно коррелирует с их эволюционным происхождением (рис. 7б). Таким образом, пять из DHN K-типа, которые демонстрируют кислый pI, являются F-DHN, в то время как все SK- и K-DHN с высокими значениями pI принадлежат к ортологичной группе Y-DHN (рис. 7b, f). Аналогичные наблюдения можно сделать и в отношении других характеристик DHN (дополнительный рис. S11), предполагая, что ортологичные группы являются лучшими индикаторами физико-химического характера DHN, чем структурные подгруппы 7 .Таким образом, в общих чертах биохимические и биофизические характеристики DHN хорошо коррелируют с тремя ортологическими группами (рис. 7g). Поскольку эти особенности, вероятно, связаны с функцией DHN, это предполагает, что функциональные исследования должны учитывать эту филогенетическую структуру, описанную здесь.
7б). Таким образом, пять из DHN K-типа, которые демонстрируют кислый pI, являются F-DHN, в то время как все SK- и K-DHN с высокими значениями pI принадлежат к ортологичной группе Y-DHN (рис. 7b, f). Аналогичные наблюдения можно сделать и в отношении других характеристик DHN (дополнительный рис. S11), предполагая, что ортологичные группы являются лучшими индикаторами физико-химического характера DHN, чем структурные подгруппы 7 .Таким образом, в общих чертах биохимические и биофизические характеристики DHN хорошо коррелируют с тремя ортологическими группами (рис. 7g). Поскольку эти особенности, вероятно, связаны с функцией DHN, это предполагает, что функциональные исследования должны учитывать эту филогенетическую структуру, описанную здесь.
Проект митохондриального генома Magnolia biondii и митохондриальная филогеномика покрытосеменных
%PDF-1.6 % 1 0 объект >поток doi:10.1371/journal.pone.0231020
 1371/journal.pone.0231020http://dx.doi.org/10.1371/journal.pone.02310202020-04-15false10.1371/journal.pone.0231020
1371/journal.pone.0231020http://dx.doi.org/10.1371/journal.pone.02310202020-04-15false10.1371/journal.pone.0231020 Новое исследование раскрывает «отвратительную тайну» Дарвина, окружающую происхождение цветковых растений — ScienceDaily
Происхождение цветковых растений, как известно, озадачило Чарльза Дарвина, описавшего их внезапное появление в летописи окаменелостей в относительно недавние геологические времена как «отвратительная тайна».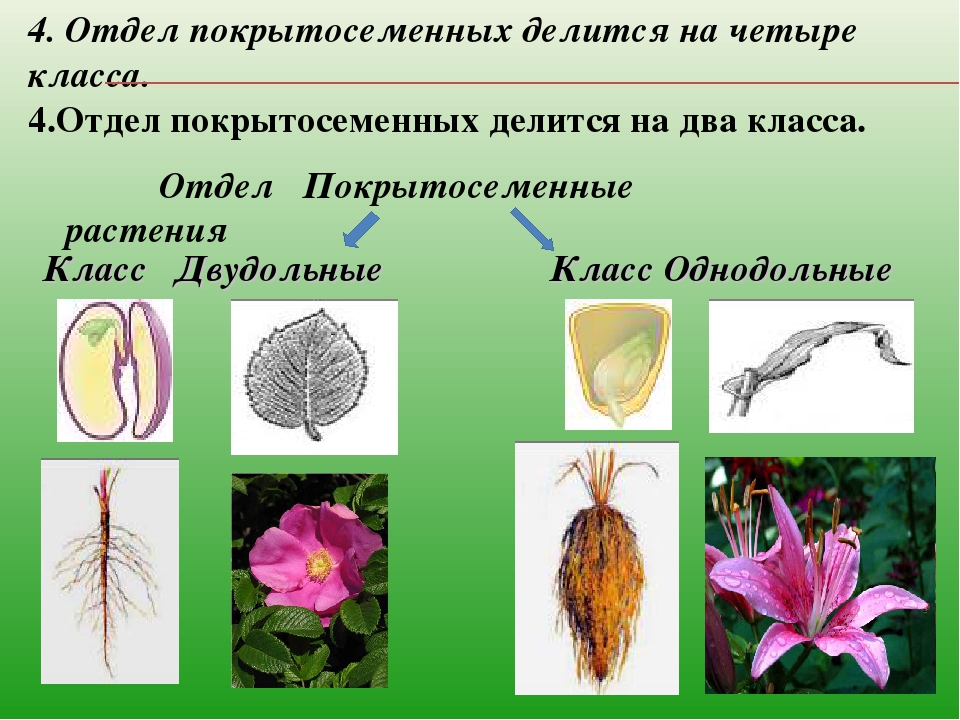 Эта тайна еще более усугубилась необъяснимым несоответствием между относительно недавней летописью окаменелостей и гораздо более древним временем происхождения цветковых растений, оцененным с использованием данных генома.
Эта тайна еще более усугубилась необъяснимым несоответствием между относительно недавней летописью окаменелостей и гораздо более древним временем происхождения цветковых растений, оцененным с использованием данных генома.
Теперь группа ученых из Швейцарии, Швеции, Великобритании и Китая, возможно, разгадала загадку. Их результаты показывают, что цветковые растения действительно возникли в юрском периоде или раньше, то есть на миллионы лет раньше, чем их самые старые бесспорные ископаемые свидетельства, согласно новому исследованию, опубликованному в научном журнале Nature Ecology & Evolution .Отсутствие более старых окаменелостей, согласно их результатам, может быть результатом низкой вероятности окаменения и редкости раннецветущих растений.
«Разнообразная группа цветковых растений очень долго жила в тени папоротников и голосеменных растений, которые доминировали в древних экосистемах. доминирующим компонентом современной фауны», — сказал ведущий автор доктор Даниэле Сильвестро из Университета Фрибурга в Швейцарии.
Цветковые растения на сегодняшний день представляют собой самую многочисленную и разнообразную группу растений в мире в современных экосистемах, намного превосходящую по численности папоротники и голосеменные растения и включающую почти все сельскохозяйственные культуры, обеспечивающие жизнедеятельность человека. Летопись окаменелостей показывает, что эта закономерность была установлена в течение последних 80-100 миллионов лет, тогда как считается, что более ранние цветковые растения были небольшими и редкими. Новые результаты показывают, что цветковые растения существовали около 100 миллионов лет, прежде чем они, наконец, стали господствовать.
«Хотя мы не ожидаем, что наше исследование положит конец спорам о происхождении покрытосеменных растений, оно дает сильную мотивацию для того, что некоторые считают охотой на снарка — цветковое растение юрского периода. «Основываясь на анализе, юрские покрытосеменные растения являются ожиданием нашей интерпретации летописи окаменелостей», — сказал соавтор, профессор Филип Донохью из Бристольского университета в Великобритании.
Выводы исследования основаны на комплексном моделировании с использованием большой глобальной базы данных о находках ископаемых, которую доктор Яову Син и его команда из Тропического ботанического сада Сишуанбаньна собрали из более чем 700 публикаций.Эти записи, насчитывающие более 15 000 человек, включали представителей многих групп растений, включая представителей пальм, орхидей, подсолнухов и гороха.
«Научные дебаты долгое время были поляризованы между палеонтологами, которые оценивают древность покрытосеменных на основе возраста самых старых окаменелостей, и молекулярными биологами, которые используют эту информацию для калибровки молекулярной эволюции по геологическому времени. Наше исследование показывает, что эти взгляды слишком упрощены; летопись окаменелостей должна быть интерпретирована», — сказала соавтор доктор Кристин Бэкон из Гетеборгского университета в Швеции.
«Буквальное прочтение летописи окаменелостей не может быть использовано для реалистичной оценки времени происхождения группы. Вместо этого нам пришлось разработать новые математические модели и использовать компьютерное моделирование для надежного решения этой проблемы».
Вместо этого нам пришлось разработать новые математические модели и использовать компьютерное моделирование для надежного решения этой проблемы».
Даже спустя 140 лет после загадки Дарвина о происхождении цветковых растений дискуссия по-прежнему занимает центральное место на научной арене. В частности, многие исследования, основанные на филогенетическом анализе современных растений и их геномов, показали, что группа возникла значительно раньше, чем указано в летописи окаменелостей, что широко оспаривается в палеонтологических исследованиях.Новое исследование, которое было основано исключительно на окаменелостях и не включало данные генома или эволюционные деревья, показывает, что более ранний возраст цветковых растений не является артефактом филогенетического анализа, но на самом деле также подтверждается палеонтологическими данными.
Соавтор, профессор Александр Антонелли, директор по науке Королевского ботанического сада в Кью, Великобритания, добавил: «Понимание того, когда цветковые растения превратились из незначительной группы в краеугольный камень большинства наземных экосистем, показывает нам, что природа динамична.
