
Половое размножение и его биологический смысл.
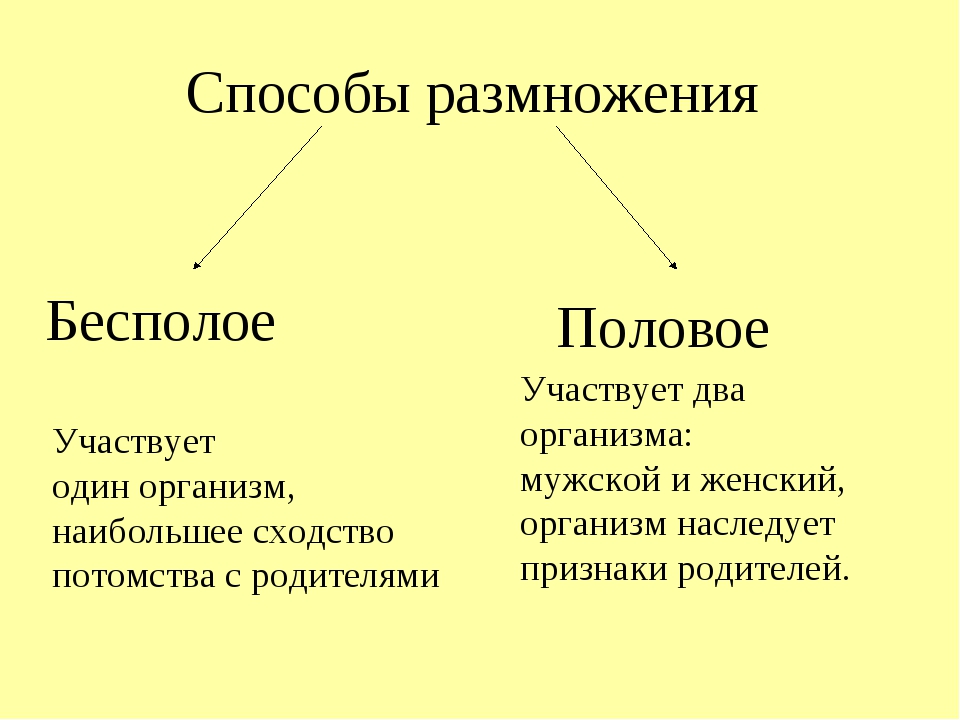
Размножение – важнейшее свойство всего живого. Вид, размножающийся только бесполым путем, может процветать достаточно длительное время, если он обитает в относительно постоянных условиях. При возникновении в среде его обитания изменений, которые вызывают гибель отдельных особей, весьма вероятно, что погибнут все особи, потому что они очень сходны генетически.
Половое размножение – более прогрессивная форма размножения, очень широко распространено в природе, как среди растений, так и среди животных. Образующиеся в процессе полового размножения организмы отличаются друг от друга генетически, а также по характеру приспособленности к условиям обитания.
При половом размножении материнским и отцовским организмами вырабатываются специализированные половые клетки – гаметы. Женские неподвижные гаметы называются яйцеклетками, мужские неподвижные – спермиями, а подвижные – сперматозоидами.
Мейоз ведет к уменьшению генетического материала вдвое, благодаря чему количество генетического материала у особей данного вида в ряду поколений остается постоянным. Во время мейоза происходит несколько важных процессов: случайное расхождение хромосом (независимое расчленение), обмен генетическим материалом между гомологичными хромосомами (кроссинговер). В результате этих процессов возникают новые комбинации генов. Поскольку ядро зиготы после оплодотворения содержит генетический материал двух родительских особей, это повышает генетическое разнообразие внутри вида. Если суть и биологическое значение полового процесса едины для всех организмов, то его формы очень разнообразны и зависят от уровня эволюционного развития, среды обитания, образа жизни и некоторых других особенностей.
Половое размножение есть у всех групп растений. Мхи растут дернинами. Мужские и женские растения оказываются рядом. Дождевая вода помогает сперматозоидам попасть на верхушки женских растений, где они сливаются с яйцеклетками, образуется зигота, из которой развивается сидящая на длинной ножке коробочка со спорами. У папоротников половые клетки развиваются на заростке, образовавшемся в результате прорастания споры. На нижней стороне заростка женские органы – архегонии, мужские – антеридии. Во влажной среде половые клетки сливаются, зигота дает начало зародышу, из которого вырастает молодой папоротник. У цветковых растений самое сложное половое размножение – двойное оплодотворение. Пыльца (мужские половые клетки) попадает на рыльце пестика (женский половой орган) и прорастает. По пыльцевой трубке спермии движутся к семязачатку. Спермии проникают в зародышевый мешок. Один сливается с яйцеклеткой и дает начало зародышу, второй спермий сливается с центральной клеткой и дает начало эндосперму – запасу питательных веществ.
У животных половое размножение связано с образованием половых клеток, которое происходит в специализированных органах – половых железах, в результате особого процесса. Половые клетки отличаются от всех остальных клеток тела уменьшенным вдвое набором хромосом. Яйцеклетка неподвижна, содержит набор питательных веществ, сперматозоиды мелкие, подвижные. Половые клетки могут образоваться в разных организмах, а могут в одном. Такие организмы называют гермафродитами (плоские черви). В природе явление гермафродитизма распространено чрезвычайно широко. Он считается самой примитивной формой полового размножения и распространен преимущественно у примитивных организмов. Одним из основных преимуществ гермафродитизма заключается в возможности самооплодотворения, что очень важно для некоторых крупных внутренних паразитов, ведущих одиночный образ жизни. Еще одной модификацией полового размножения является партеногенез. При таком способе размножения женская гамета развивается в дочернюю особь без оплодотворения мужской гаметой.
Половое размножение имеет очень большие эволюционные преимущества по сравнению с бесполым. Сущность полового размножения заключается в объединении в наследственном материале потомка генетической информации из двух разных источников – родителей. Оплодотворение у животных может быть наружным или внутренним. При слиянии образуется зигота с двойным набором хромосом.
В ядре зиготы все хромосомы становятся парными: в каждой паре одна из хромосом отцовская, другая – материнская. Дочерний организм, который разовьется из такой зиготы, в одинаковой мере снабжен наследственной информацией обоих родителей.
Биологический смысл полового размножения состоит в том, что возникающие организмы могут сочетать полезные признаки отца и матери. Такие организмы более жизнеспособны. Половое размножение играет важную роль в эволюции организмов.
06 | ЮУГМУ, Челябинск
Практическое занятие №3
ТЕМА: РАЗМНОЖЕНИЕ НА ОРГАНИЗМЕННОМ УРОВНЕ. СПОСОБЫ
СПОСОБЫ
БЕСПОЛОГО И ПОЛОВОГО РАЗМНОЖЕНИЯ
Цели занятия:
1) Уяснить биологическую сущность и преимущества полового размножения над бесполым.
2) Изучить особенности гаметогенеза в связи со специфическими функциями гамет.
3) Обратить внимание на влияние никотина, алкоголя и наркотиков на наследственность человека.
Базисные знания:
1) Из курса биологии средней школы Вы должны знать основные закономерности мейоза и фазы гаметогенеза.
Учебная карта занятия:
А) Вопросы для подготовки к занятию:
- Размножение – универсальное свойство, обеспечивающее материальную непрерывность живого.
- Эволюция размножения:
а) бесполое;
б) половое;
в) партеногенез и его разновидности.
- Половой процесс как механизм обмена наследственной информацией внутри вида.

- Цитологические механизмы размножения (мейоз и его цитологическая характеристика).
- Закономерности овогенеза и сперматогенеза. Строение половых клеток. Типы яйцеклеток.
- Влияние никотина, алкоголя и наркотиков на наследственность человека.
Б) Список основной и дополнительной литературы для подготовки к занятию приведен в конце тетради на странице 92.
В) Задания для учебно-исследовательской работы обучающихся:
Задание 1. Изучение микроскопического строения семенника крысы.
Рассмотреть на большом увеличении микроскопа препарат “Поперечный срез семенника крысы”. Зарисовать строение семенного канальца, отметив на рисунке:
- сперматогонии (круглые клетки с темными ядрами, расположенные в основании стенки канальцев);
- сперматоциты 1-ого и 2-ого порядков (расположены ближе к просвету канальца, обладают более светлыми, крупными ядрами и имеют более крупные размеры) ;
3. сперматозоиды (имеют вид тонких нитей, расположенных в просвете канальца).
сперматозоиды (имеют вид тонких нитей, расположенных в просвете канальца).
|
1.
2.
3. |
Задание 2. Изучение микроскопического строения яичника млекопитающего.
Рассмотреть на малом увеличении микроскопа микропрепарат “Срез яичника млекопитающего”.
Зарисовать зрелый фолликул – граафов пузырек, отметив:
1. стенку фолликула;
2. полость фолликула;
3. яйценосный бугорок (выступ, образованный фолликулярными клетками)
4. яйцеклетку (ооцит 2-ого порядка, расположенный внутри яйценосного бугорка).
|
1.
2.
3.
4.
|
Г) Практические навыки, которыми должен овладеть обучающийся по теме занятия:
1) Определение разных стадий сперматогенеза на срезе семенника крысы.
2) Идентификация зрелых фолликулов на срезе яичника кошки.
Подпись преподавателя: _________________________________________________________
§ 25. Размножение организмов. Биология 9 класс Пасечник
1. Все ли живые существа способны к размножению?
Все живые существа способны размножаться. Это важнейшее свойство живого.
2. Какой вид размножения появился первым?
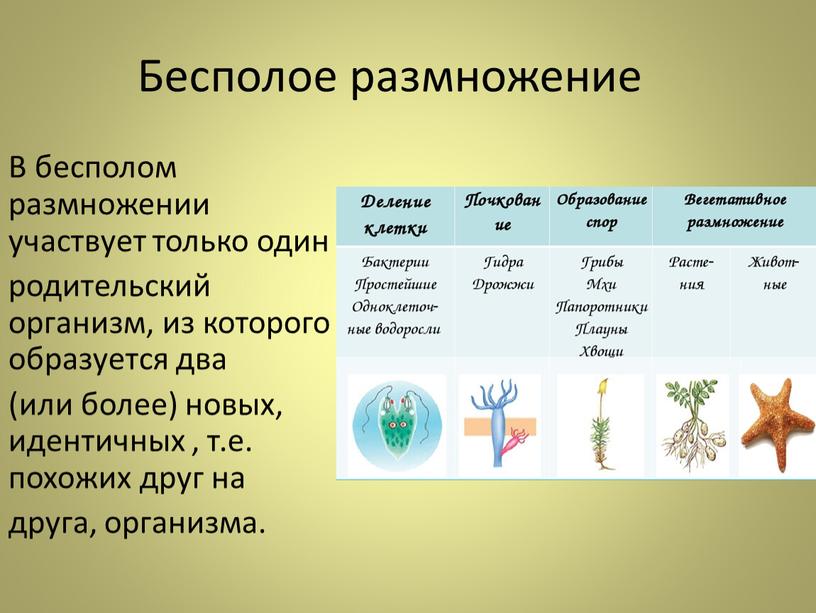
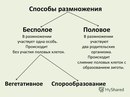
Древнейшим способом размножения на Земле было бесполое размножение. При бесполом размножении одна или несколько клеток тела родительской особи делятся. При этом образуется одна или несколько дочерних особей, во всём схожих с родительской.
При бесполом размножении одна или несколько клеток тела родительской особи делятся. При этом образуется одна или несколько дочерних особей, во всём схожих с родительской.
3. У каких простейших появился половой процесс?
Уже у простейших (например, инфузория) встречается половой процесс («конъюгация» — лат. «соединение»), первоначально не имеющий прямого отношения к размножению. В ходе полового процесса особи обмениваются между собой генетической информацией, что приводит к их взаимному обновлению и повышает их выживаемость.
В ходе эволюции соединение полового процесса и размножения привело к возникновению полового размножения.
4. Какие растения называются двудомными?
Двудомные растения — виды растений, у которых мужские (тычиночные) и женские (пестичные) цветки находятся не на одной особи, а на разных. Например, ива, тополь и др.
Вопросы
1. Какие виды бесполого размножения вы можете назвать?
Какие виды бесполого размножения вы можете назвать?
Среди форм бесполого размножения различают деление клетки, почкование, делением тела, спорами, вегетативное.
В любом случае для размножения достаточно одной особи, в результате образуются особи, идентичные материнской.
2. Почему гидры размножаются бесполым путём только в благоприятных условиях?
После того, как маленькая гидра отделяется от материнского организма, она обычно располагается рядом с материнским организмом. «Мать» и «дочь» начинают конкурировать за пищу. Вот почему гидры размножаются почкованием только тогда, когда условия существования хорошие и пищи достаточно.
3. Перечислите способы вегетативного размножения у высших растений.
Растения могут размножаться стеблем или его частью и видоизменениями: отводками (смородина), черенками (тополь), усами (земляника), клубнями (картофель), корневищами (ирис), луковицами (лук, чеснок, тюльпан). Возможно также вегетативное размножение корнями (малина, слива) и корнеклубнями (георгин). В определённых условиях растение может размножаться и черенком листа (бегония)
Возможно также вегетативное размножение корнями (малина, слива) и корнеклубнями (георгин). В определённых условиях растение может размножаться и черенком листа (бегония)
4. В чём недостаток бесполого размножения?
При таком способе размножения все потомки имеют абсолютно такой же генотип, как и родительская особь: ведь они развиваются из клеток тела этой особи. Следовательно, при бесполом размножении не происходит увеличения генетического разнообразия, которое может оказаться очень полезным при изменении условий существования вида.
5. В чём преимущество полового размножения перед бесполым?
При половом размножении генетическое разнообразие в потомстве увеличивается, а значит, растёт и жизнеспособность по сравнению с родительской.
6. Каких животных называют гермафродитами?
Гермафродиты — обоеполые организмы, способные производить одновременно два вида половых клеток — как мужские, так и женские. Это некоторые кишечнополостные, все плоские черви, отдельные виды кольчатых червей, а также некоторые моллюски и даже рыбы и ящерицы.
Это некоторые кишечнополостные, все плоские черви, отдельные виды кольчатых червей, а также некоторые моллюски и даже рыбы и ящерицы.
7. Как устроены сперматозоид и яйцеклетка?
Яйцеклетка неподвижна, обычно крупных размеров и содержит запасы питательных веществ. Она обеспечивает зародыш будущего организма основной массой цитоплазмы и, особенно у яйцекладущих животных, питательным материалом (желтком). Яйцеклетка в сотни раз, а у животных с богатыми желтком яйцеклетками — в миллионы раз больше сперматозоида. Яйцеклетка большинства животных окружена защитными оболочками.
Сперматозоиды очень малы и подвижны. У млекопитающих сперматозоид состоит из головки (её длина около 5-10 мкм), шейки и хвостика (их общая длина около 60 мкм). В головке расположено ядро, содержащее гаплоидный набор хромосом. Цитоплазмы в головке очень мало. В шейке находятся небольшое число митохондрий, вырабатывающих энергию для движения сперматозоида, и центриоль, обеспечивающая колебания жгутика, лежащего вдоль оси хвостика.
Задания
Сравните процессы бесполого и полового размножения. Объясните, что общего и чём различия этих процессов. В чём заключается их биологическая сущность?
Общее у этих процессов то, что в любом случае количество особей увеличивается.
Сравнительная характеристика бесполого и полового размножения, их сущность и биологическое значение
1. Сравнительная характеристика бесполого и полового размножения, их сущность и биологическое значение
2. Типы полового размножения
ПоловоеПринципиальное отличие полового размножения от бесполого
состоит в том, что в нем участвуют обычно два
родительских организма, признаки которых
перекомбинируются у потомства. Половое
размножение свойственно всем эукариотам, но
преобладает оно у животных и высших растений.
Переход к этому типу размножения имел огромное значение
для эволюции жизни на Земле.
 Половое размножение
Половое размножениесоздает бесконечное разнообразие особей, в том
числе и таких, которые успешно адаптируются к
изменчивым внешним условиям, «завоевывают мир»,
распространяясь в новые места обитания, и оставляют
потомство, передавая ему свой наследственный
материал. Потомки же двух успешных родительских
особей могут оказаться обладателями еще более
удачной комбинации наследственных признаков, и
соответственно они разовьют успех родителей. Особи с
неудачной комбинацией признаков будут
элиминированы естественным отбором. Таким
образом, половое размножение создает богатый
материал для естественного отбора и эволюции.
Любопытно и другое: само возникновение особи как
индивидуальности, неделимого и смертного существа,
является результатом перехода к половому
размножению. При бесполом размножении клетка
бесконечно делится, повторяя саму себя: она
потенциально бессмертна, но особью может быть
названа только условно, так как не отличимо от
неопределенного множества дочерних клеток.
 При
Приполовом размножении, напротив, все потомки
различаются между собой и отличаются от родителей, а
те с течением времени умирают, унося с собой
свойственные им неповторимые особенности.
Американский зоолог Р.Хегнер, обсуждая простейших,
выразил это таким образом: «Они приобрели очередное
новшество — пол; цена этого приобретения — неминуемая
естественная гибель… Не велика ли эта цена?»[
Гормональная регуляция размножения у
млекопитающих. М., 1987] Подчеркнем однако, что
одновременно открылись возможности для развития и
совершенствования, и они привели к появлению
разнообразных живых форм, не сопоставимых по
уровню организации с теми организмами, которые
остановились на бесполом размножении.
Бесполое
Бесполое размножение, или агамогенез форма размножения, при которой
организм воспроизводит себя
самостоятельно, без всякого участия другой
особи. Следует отличать бесполое
размножение от однополого размножения
(партеногенеза), который является особой
формой полового размножения.

Крайняя простота этого способа размножения,
связанная с относительной простотой
организации одноклеточных организмов,
позволяет размножаться очень быстро. Так, в
благоприятных условиях количество бактерий
может удваиваться каждые 30-60 минут.
Размножающийся бесполым путем
организм способен бесконечно
воспроизводить себя, пока не произойдет
спонтанное изменение генетического
материала — мутация. Если эта мутация
благоприятна, она сохранится в потомстве
мутировавшей клетки, которое будет
представлять собой новый клеточный клон.
Бесполое размножение, воспроизводящее
идентичные исходному организму особи, не
способствует появлению организмов с
новыми вариантами признаков, а тем
самым ограничивает возможность
приспособления видов к новым для них
условиям среды. Средством преодоления
этой ограниченности стал переход к
половому размножению.
3. Бесполое размножение
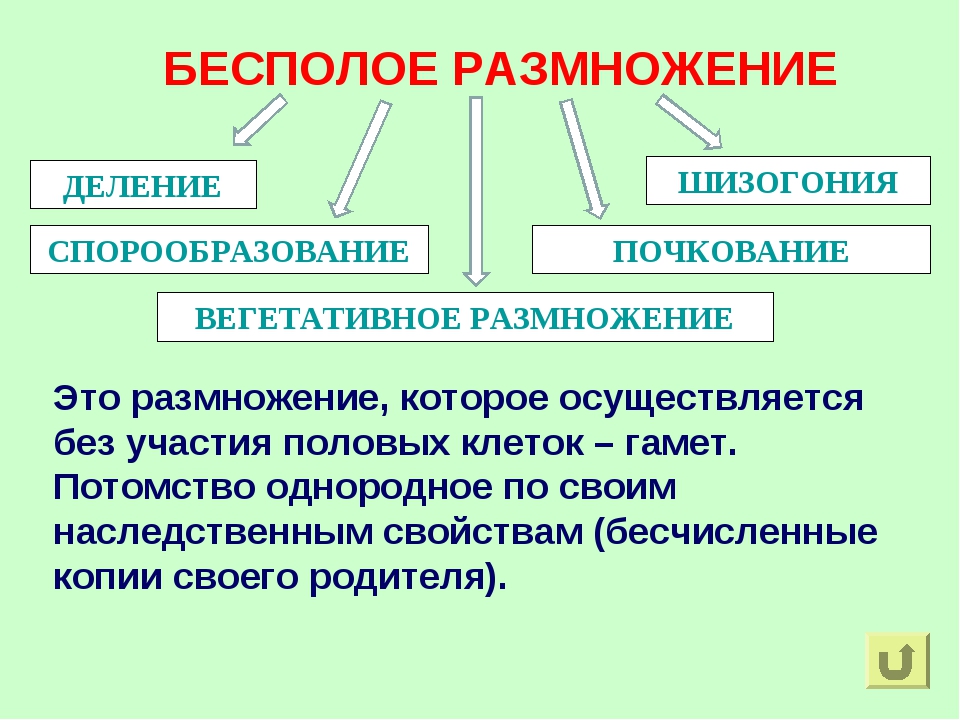
Различают следующие способы бесполого размножения: деление, почкование, фрагментация,полиэмбриония, спорообразование, вегетативное размножение.

Деление — способ бесполого размножения, характерный для одноклеточных организмов, при котором
материнская особь делится на две или большее количество дочерних клеток. Можно выделить: а) простое
бинарное деление (прокариоты), б) митотическое бинарное деление (простейшие, одноклеточные
водоросли), в) множественное деление, или шизогонию (малярийный плазмодий, трипаносомы). Во время
деления парамеции (1) микронуклеус делится митозом, макронуклеус — амитозом. Во время шизогонии
(2) сперва многократно митозом делится ядро, затем каждое из дочерних ядер окружается цитоплазмой, и
формируются несколько самостоятельных организмов.
Почкование — способ бесполого размножения, при котором новые особи образуются в виде выростов на
теле родительской особи (3). Дочерние особи могут отделяться от материнской и переходить к
самостоятельному образу жизни (гидра, дрожжи), могут остаться прикрепленными к ней, образуя в этом
случае колонии (коралловые полипы).

Фрагментация (4) — способ бесполого размножения, при котором новые особи образуются из
фрагментов (частей), на которые распадается материнская особь (кольчатые черви, морские звезды,
спирогира, элодея). В основе фрагментации лежит способность организмов к регенерации.
Полиэмбриония — способ бесполого размножения, при котором новые особи образуются из фрагментов
(частей), на которые распадается эмбрион (монозиготные близнецы).
Вегетативное размножение — способ бесполого размножения, при котором новые особи образуются или
из частей вегетативного тела материнской особи, или из особых структур (корневище, клубень и др.),
специально предназначенных для этой формы размножения. Вегетативное размножение характерно для
многих групп растений, используется в садоводстве, огородничестве, селекции растений (искусственное
вегетативное размножение).
Спорообразование (6) — размножение посредством спор. Споры — специализированные клетки, у
большинства видов образуются в особых органах — спорангиях.
 У высших растений образованию спор
У высших растений образованию спорпредшествует мейоз.
Клонирование — комплекс методов, используемых человеком для получения генетически идентичных копий
клеток или особей.Клон — совокупность клеток или особей, произошедших от общего предка путем
бесполого размножения. В основе получения клона лежит митоз (у бактерий — простое деление).
4. Половое размножение
осуществляется при участии двух родительских особей (мужской и женской), укоторых в особых органах образуются специализированные клетки — гаметы. Процесс
формирования гамет называется гаметогенезом, основным этапом гаметогенеза является мейоз.
Дочернее поколение развивается из зиготы — клетки, образовавшейся в результате слияния
мужской и женской гамет. Процесс слияния мужской и женской гамет
называется оплодотворением. Обязательным следствием полового размножения является
перекомбинация генетического материала у дочернего поколения.
В зависимости от особенностей строения гамет, можно выделить следующие формы полового
размножения: изогамию, гетерогамию и овогамию.
Изогамия (1) — форма полового размножения, при которой гаметы (условно женские и условно
мужские) являются подвижными и имеют одинаковые морфологию и размеры.
Гетерогамия (2) — форма полового размножения, при которой женские и мужские гаметы
являются подвижными, но женские — крупнее мужских и менее подвижны.
Овогамия (3) — форма полового размножения, при которой женские гаметы неподвижные и более
крупные, чем мужские гаметы. В этом случае женские гаметы называются яйцеклетками, мужские
гаметы, если имеют жгутики, — сперматозоидами, если не имеют, — спермиями.
Овогамия характерна для большинства видов животных и растений. Изогамия и гетерогамия
встречаются у некоторых примитивных организмов (водоросли). Кроме вышеперечисленных, у
некоторых водорослей и грибов имеются формы размножения, при которых половые клетки не
образуются: хологамия и конъюгация. При хологамии происходит слияние друг с другом
одноклеточных гаплоидных организмов, которые в данном случае выступают в роли гамет.

Образовавшаяся диплоидная зигота затем делится мейозом с образованием четырех гаплоидных
организмов. При конъюгации (4) происходит слияние содержимого отдельных гаплоидных клеток
нитевидных талломов. По специально образующимся каналам содержимое одной клетки
перетекает в другую, образуется диплоидная зигота, которая обычно после периода покоя также
делится мейозом.
Бесполое
Участвует два организма
Участвует один организм
Участвуют половые клетки
(гаметы), полученные путем
мейоза
Участвуют соматические клетки,
размножающиеся митозом.
Дети получаются разные
(происходит перекомбинация
признаков отца и матери,
повышается генетическое
разнообразие популяции)
Дети получаются одинаковые,
копии родителя (в сельском
хозяйстве – позволяет быстро
увеличить численность
организмов, сохраняя все
признаки сорта)
Методы размножения – Биология
OpenStaxCollege
[латексная страница]
Цели обучения
К концу этого раздела вы сможете:
- Описать преимущества и недостатки бесполого и полового размножения
- Обсудить методы бесполого размножения
- Обсудить методы полового размножения
Животные производят потомство путем бесполого и/или полового размножения. Оба метода имеют преимущества и недостатки.Бесполое размножение дает потомство, генетически идентичное родителю, потому что все потомки являются клонами исходного родителя. Одна особь может произвести потомство бесполым путем, и большое количество потомства может быть произведено быстро. В стабильной или предсказуемой среде бесполое размножение является эффективным средством размножения, поскольку все потомство будет адаптировано к этой среде. В нестабильной или непредсказуемой среде виды, размножающиеся бесполым путем, могут оказаться в невыгодном положении, потому что все потомство генетически идентично и может не иметь генетических вариаций, необходимых для выживания в новых или иных условиях.С другой стороны, высокие темпы бесполого размножения могут позволить быстро реагировать на изменения окружающей среды, если у людей есть мутации. Дополнительным преимуществом бесполого размножения является то, что колонизация новых мест обитания может быть проще, когда человеку не нужно искать себе пару для размножения.
Оба метода имеют преимущества и недостатки.Бесполое размножение дает потомство, генетически идентичное родителю, потому что все потомки являются клонами исходного родителя. Одна особь может произвести потомство бесполым путем, и большое количество потомства может быть произведено быстро. В стабильной или предсказуемой среде бесполое размножение является эффективным средством размножения, поскольку все потомство будет адаптировано к этой среде. В нестабильной или непредсказуемой среде виды, размножающиеся бесполым путем, могут оказаться в невыгодном положении, потому что все потомство генетически идентично и может не иметь генетических вариаций, необходимых для выживания в новых или иных условиях.С другой стороны, высокие темпы бесполого размножения могут позволить быстро реагировать на изменения окружающей среды, если у людей есть мутации. Дополнительным преимуществом бесполого размножения является то, что колонизация новых мест обитания может быть проще, когда человеку не нужно искать себе пару для размножения.
Во время полового размножения генетический материал двух особей объединяется для получения генетически разнообразного потомства, отличающегося от своих родителей. Считается, что генетическое разнообразие потомства, производимого половым путем, дает видам больше шансов выжить в непредсказуемой или меняющейся среде.Виды, размножающиеся половым путем, должны поддерживать два разных типа особей, самцов и самок, что может ограничить способность колонизировать новые места обитания, поскольку должны присутствовать представители обоих полов.
Бесполое размножение встречается у прокариотических микроорганизмов (бактерий) и у некоторых эукариотических одноклеточных и многоклеточных организмов. Существует несколько способов бесполого размножения животных.
Деление
Деление, также называемое бинарным делением, происходит у прокариотических микроорганизмов и у некоторых беспозвоночных многоклеточных организмов.После периода роста организм разделяется на два отдельных организма.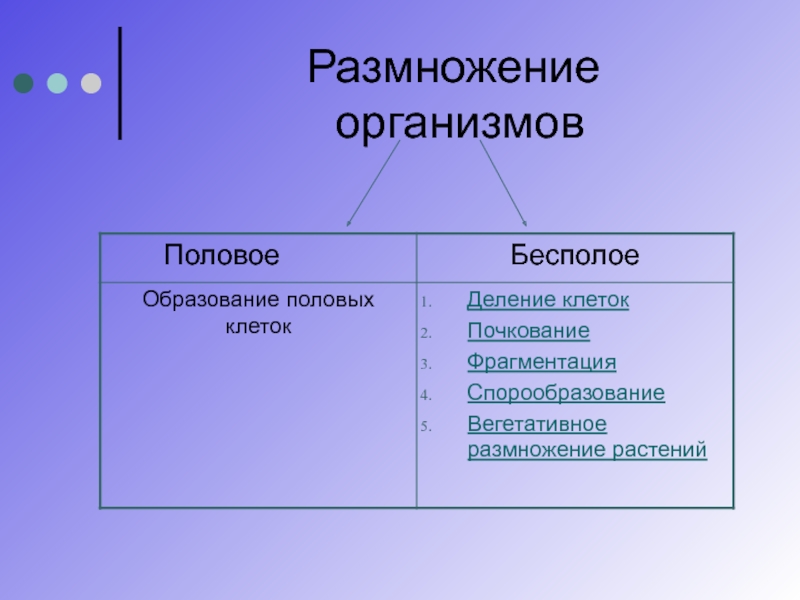 Некоторые одноклеточные эукариотические организмы подвергаются бинарному делению путем митоза. У других организмов часть особи обособляется и образует вторую особь. Этот процесс происходит, например, у многих астероидных иглокожих путем расщепления центрального диска. Некоторые морские анемоны и некоторые коралловые полипы ([ссылка]) также размножаются делением.
Некоторые одноклеточные эукариотические организмы подвергаются бинарному делению путем митоза. У других организмов часть особи обособляется и образует вторую особь. Этот процесс происходит, например, у многих астероидных иглокожих путем расщепления центрального диска. Некоторые морские анемоны и некоторые коралловые полипы ([ссылка]) также размножаются делением.
Коралловые полипы размножаются бесполым путем делением. (кредит: Г. П. Шмаль, менеджер NOAA FGBNMS)
Бутонизация
Почкование — это форма бесполого размножения, возникающая в результате разрастания части клетки или участка тела, что приводит к разделению исходного организма на две особи.Почкование обычно происходит у некоторых беспозвоночных животных, таких как кораллы и гидры. У гидр формируется почка, которая развивается во взрослую особь и отрывается от основного тела, как показано на [ссылка], тогда как при почковании кораллов почка не отделяется и размножается как часть новой колонии.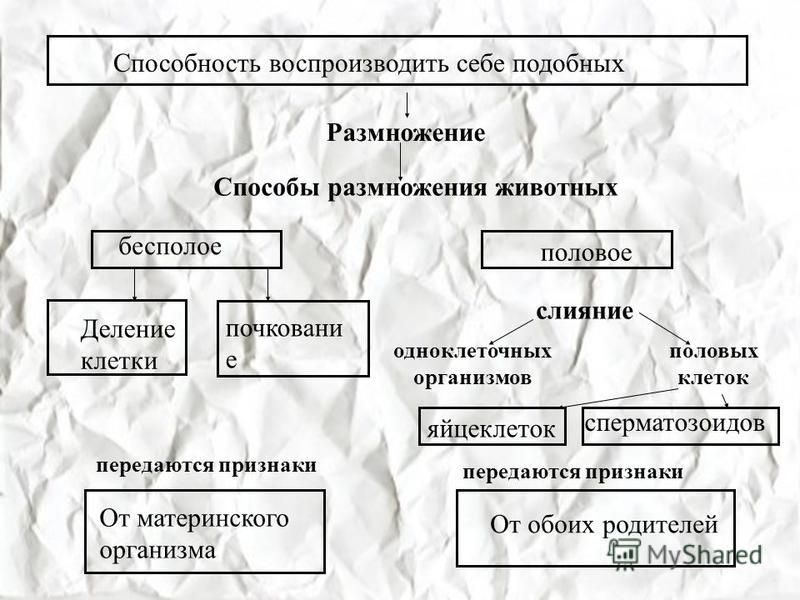
Гидры размножаются бесполым путем путем почкования.
Ссылка на обучение
Посмотрите видео почкования гидры.
Фрагментация
Фрагментация — это разделение тела на две части с последующей регенерацией.Если животное способно к фрагментации, а часть достаточно велика, вырастет отдельная особь.
Например, у многих морских звезд бесполое размножение осуществляется путем фрагментации. [ссылка] иллюстрирует морскую звезду, у которой сломана рука человека и восстанавливается новая морская звезда. Известно, что работники рыболовства пытались убить морских звезд, поедающих их моллюсков или устриц, разрезая их пополам и бросая обратно в океан. К несчастью для рабочих, каждая из двух частей может регенерировать новую половину, в результате чего вдвое больше морских звезд охотятся на устриц и моллюсков.Фрагментация также происходит у кольчатых червей, турбеллярий и порифер.
Морские звезды могут размножаться путем фрагментации.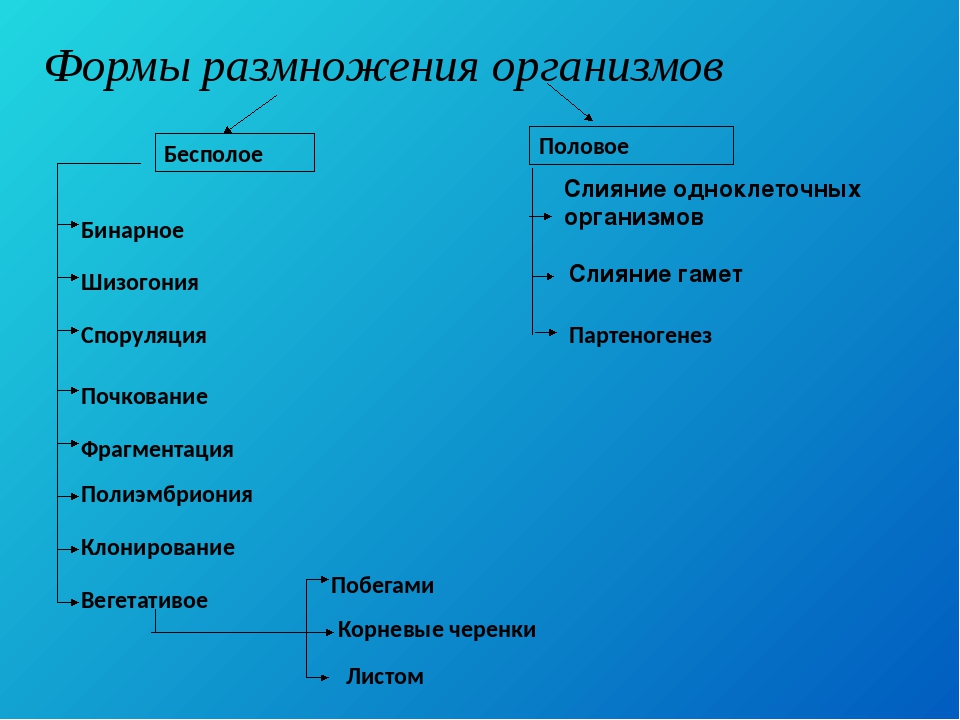 Большая рука, фрагмент другой морской звезды, превращается в новую особь.
Большая рука, фрагмент другой морской звезды, превращается в новую особь.
Обратите внимание, что при фрагментации обычно наблюдается заметная разница в размерах особей, тогда как при делении образуются две особи приблизительного размера.
Партеногенез
Партеногенез — форма бесполого размножения, при которой яйцеклетка развивается в полноценную особь без оплодотворения.Полученное потомство может быть либо гаплоидным, либо диплоидным, в зависимости от процесса и вида. Партеногенез происходит у беспозвоночных, таких как водоплавающие, коловратки, тли, палочники, некоторые муравьи, осы и пчелы. Пчелы используют партеногенез для производства гаплоидных самцов (трутней). При оплодотворении яиц развиваются диплоидные самки, а при кормлении оплодотворенных яиц специальным кормом (так называемым маточным молочком) образуется матка.
Некоторые позвоночные животные, такие как некоторые рептилии, амфибии и рыбы, также размножаются посредством партеногенеза. Хотя партеногенез чаще встречается у растений, он наблюдался у видов животных, которые были разделены по полу в наземных или морских зоопарках. Две самки дракона Комодо, акула-молот и чернотопая акула произвели партеногенных детенышей, когда самок изолировали от самцов.
Хотя партеногенез чаще встречается у растений, он наблюдался у видов животных, которые были разделены по полу в наземных или морских зоопарках. Две самки дракона Комодо, акула-молот и чернотопая акула произвели партеногенных детенышей, когда самок изолировали от самцов.
Половое размножение — это объединение (обычно гаплоидных) репродуктивных клеток двух особей с образованием третьего (обычно диплоидного) уникального потомства. Половое размножение дает потомство с новыми комбинациями генов.Это может быть адаптивным преимуществом в нестабильной или непредсказуемой среде. Как люди, мы привыкли думать, что животные имеют два разных пола — мужской и женский — определенные при зачатии. Однако в животном мире существует множество вариаций на эту тему.
Гермафродитизм
Гермафродитизм возникает у животных, у которых одна особь имеет как мужские, так и женские репродуктивные органы. Беспозвоночные, такие как дождевые черви, слизни, ленточные черви и улитки, показанные на [ссылка], часто являются гермафродитами. Гермафродиты могут самооплодотворяться или спариваться с другим представителем своего вида, оплодотворяя друг друга и производя потомство. Самооплодотворение распространено у животных с ограниченной подвижностью или неподвижных, таких как ракушки и моллюски.
Гермафродиты могут самооплодотворяться или спариваться с другим представителем своего вида, оплодотворяя друг друга и производя потомство. Самооплодотворение распространено у животных с ограниченной подвижностью или неподвижных, таких как ракушки и моллюски.
Многие улитки гермафродиты. Когда две особи спариваются, каждая из них может произвести до ста яиц. (кредит: Ассаф Штильман)
Определение пола у млекопитающих определяется генетически наличием X- и Y-хромосом.Лица, гомозиготные по X (XX), — женщины, а гетерозиготные (XY) — мужчины. Наличие Y-хромосомы вызывает развитие мужских признаков, а ее отсутствие приводит к женским признакам. Система XY также встречается у некоторых насекомых и растений.
Определение пола птиц зависит от наличия Z- и W-хромосом. Гомозиготность по Z (ZZ) дает мальчика, а гетерозигота (ZW) дает женщину. W, по-видимому, играет важную роль в определении пола человека, подобно Y-хромосоме у млекопитающих.Эту систему используют некоторые рыбы, ракообразные, насекомые (например, бабочки и мотыльки) и рептилии.
Пол некоторых видов определяется не генетикой, а некоторыми аспектами окружающей среды. Определение пола у некоторых крокодилов и черепах, например, часто зависит от температуры во время критических периодов развития яиц. Это называется определением пола в окружающей среде или, более конкретно, определением пола в зависимости от температуры. У многих черепах при более низких температурах во время инкубации яиц появляются самцы, а при более высоких температурах — самки.У некоторых крокодилов при умеренной температуре появляются самцы, а при тепле и холоде — самки. У некоторых видов пол зависит как от генетики, так и от температуры.
Особи некоторых видов в течение жизни меняют пол, чередуя самцов и самок. Если человек в первую очередь женщина, это называется протогинией или «первой женщиной», если это мужчина в первую очередь, это называется протандрией или «первым мужчиной». Устрицы, например, рождаются самцами, растут, становятся самками и откладывают яйца; некоторые виды устриц меняют пол несколько раз.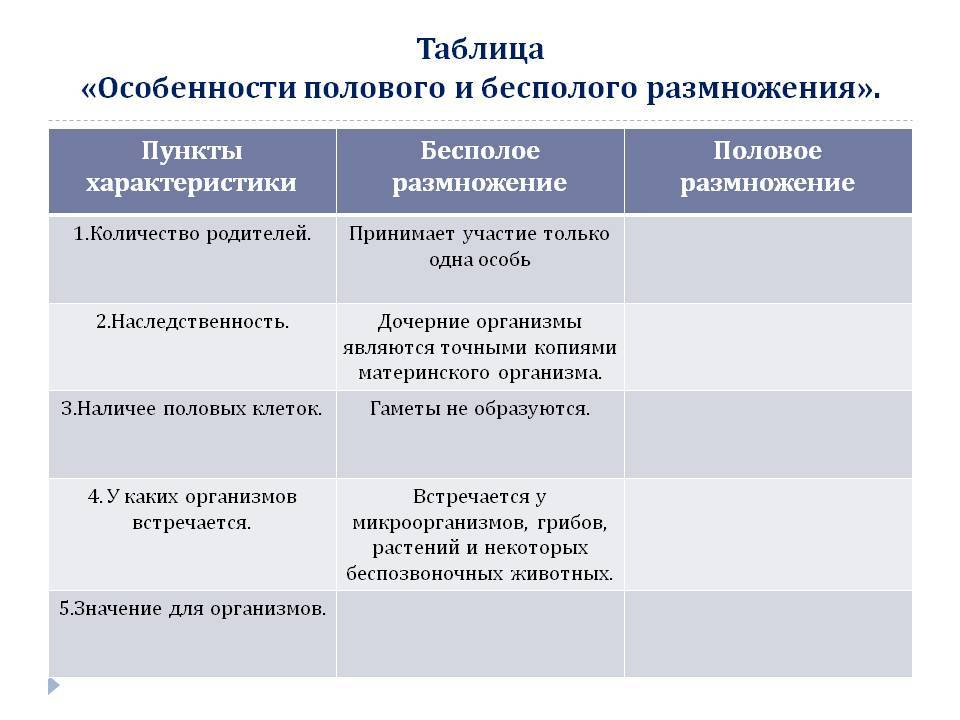
Размножение может быть бесполым, когда одна особь производит генетически идентичное потомство, или половым, когда генетический материал двух особей объединяется для получения генетически разнообразного потомства. Бесполое размножение происходит путем деления, почкования и фрагментации. Половое размножение может означать соединение сперматозоидов и яйцеклеток в телах животных или может означать выброс сперматозоидов и яйцеклеток в окружающую среду. Человек может быть одного пола или обоих полов; он может начать как один пол и измениться в течение своей жизни, или он может остаться мужчиной или женщиной.
Какая форма воспроизводства считается лучшей в стабильной среде?
- бесполое
- сексуальный
- бутонизация
- партеногенез
Какая форма репродукции может возникнуть в результате повреждения исходного животного?
- бесполое
- фрагментация
- бутонизация
- партеногенез
Какая форма размножения полезна для малоподвижных животных, размножающихся половым путем?
- деление
- бутонизация
- партеногенез
- гермафродитизм
Генетически уникальные особи производятся посредством ________.
- половое размножение
- партеногенез
- бутонизация
- фрагментация
Почему полезно половое размножение, если только половина животных может произвести потомство и две отдельные клетки должны быть объединены, чтобы образовать третью?
Половое размножение дает новую комбинацию генов в потомстве, которая может лучше помочь им пережить изменения в окружающей среде и помочь в выживании вида.
Что определяет пол потомков птиц и млекопитающих?
Наличие W-хромосомы у птиц определяет женский пол, а наличие Y-хромосомы у млекопитающих определяет мужской пол.Отсутствие этих хромосом и однородность потомства (ZZ или XX) приводит к развитию другого пола.
Глоссарий
- бесполое размножение
- форма воспроизводства, которая дает потомство, генетически идентичное родителю
- бутонизация
- форма бесполого размножения, возникающая в результате разрастания части клетки, что приводит к разделению исходного животного на две особи
- деление
- (также бинарное деление) метод, при котором многоклеточные организмы увеличиваются в размерах, или бесполое размножение, при котором одноклеточный организм делится на два отдельных организма путем митоза
- фрагментация
- разрезание или фрагментация исходного животного на части и выращивание отдельного животного из каждой части
- гермафродитизм
- состояние наличия как мужских, так и женских половых органов у одного и того же человека
- партеногенез
- форма бесполого размножения, при которой яйцеклетка развивается в полную особь без оплодотворения
- половое размножение
- смешение генетического материала двух особей для получения генетически уникального потомства
влияние бесполого и полового размножения на пространственную генетическую структуру внутри и между популяциями двудомного растения Marchantia inflexa (Marchantiaceae) | Анналы ботаники
Аннотация
Исходная информация и цели
У раздельнополых растений половое размножение требует непосредственной близости к потенциальным партнерам, но клональный рост может увеличить это расстояние и, следовательно, снизить вероятность спаривания. Сокращение половых отростков может привести к уменьшению расселения и потока генов между популяциями. На поток генов и рост клонов может дополнительно влиять размер участка среды обитания. Влияние размера среды обитания и способа размножения (половое или бесполое размножение) на пространственную генетическую структуру и сегрегацию полов было проверено путем количественного определения распределения генотипов и полов с использованием раздельнополой печеночницы Marchantia inflexa.
Сокращение половых отростков может привести к уменьшению расселения и потока генов между популяциями. На поток генов и рост клонов может дополнительно влиять размер участка среды обитания. Влияние размера среды обитания и способа размножения (половое или бесполое размножение) на пространственную генетическую структуру и сегрегацию полов было проверено путем количественного определения распределения генотипов и полов с использованием раздельнополой печеночницы Marchantia inflexa.
Методы
Образцы растений были отобраны из пяти пар малых и больших участков местообитаний для определения внутрипопуляционной и межпопуляционной пространственной генетической структуры с использованием 12 микросателлитных маркеров.Пространственное распределение рассчитывалось как вероятность того, что пары особей были одного пола или генотипа, и определялось, как на эту вероятность влиял размер участка среды обитания (маленький/большой).
Ключевые результаты
В популяциях преобладает бесполое размножение, и бесполое расселение также имело место среди популяций.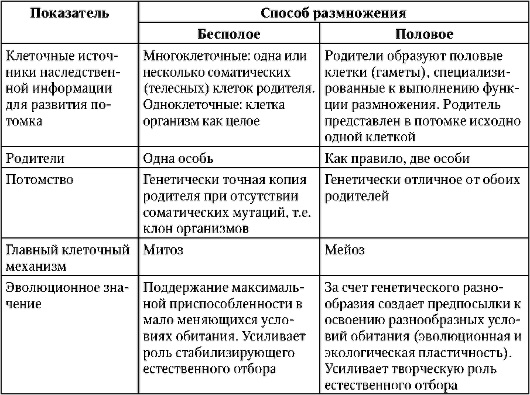 Внутри популяций наблюдалась пространственная сегрегация полов; мужчины чаще находились рядом с особями того же пола, чем женщины. Хотя вероятность того, что представители обоих полов будут рядом с представителями одного пола, была одинаково выше на небольших участках среды обитания, на больших участках среды обитания вероятность того, что мужские генотипы будут рядом с клонами, почти на 15 % выше, чем у женских генотипов.
Внутри популяций наблюдалась пространственная сегрегация полов; мужчины чаще находились рядом с особями того же пола, чем женщины. Хотя вероятность того, что представители обоих полов будут рядом с представителями одного пола, была одинаково выше на небольших участках среды обитания, на больших участках среды обитания вероятность того, что мужские генотипы будут рядом с клонами, почти на 15 % выше, чем у женских генотипов.
Выводы
Результаты показывают половые различия в скоплении клонов, которые зависели от размера среды обитания, что свидетельствует о различной колонизации и/или выживании между самцами и самками. Различная структура полов и генотипов внутри и между популяциями имеет значение для устойчивости популяций и взаимодействия между ними. Это исследование демонстрирует, что изучение только полов, а не их генотипов (или наоборот) может ограничить наше понимание того, в какой степени репродуктивные способы (половые или бесполые) влияют на генетическую структуру как внутри популяций, так и между ними.
ВВЕДЕНИЕ
Многие виды растений используют как клональное, так и половое размножение, при этом клональное размножение позволяет растениям поддерживать и распространять свои популяции, а половое размножение обеспечивает преимущества полового процесса, особенно эволюционный потенциал (Frankham, 1995). Несмотря на это преимущество клонального размножения, длительные периоды клонального роста могут вызывать как пространственное разделение полов (SSS), так и пространственную генетическую структуру (SGS) внутри популяций, что приводит к сокращению или полной потере полового размножения (Barrett, 2015).По мере того, как родительские генотипы или генеты увеличиваются в размерах за счет клональной экспансии, увеличивается их близость к особи другого пола или другого генотипа. Это увеличение расстояния может усилить инбридинг среди клонов или раметов у самосовместимых видов (Eckert, 2000; Raabová et al. , 2015) и может ограничить половое размножение у облигатных ауткроссинговых видов, таких как самонесовместимые и раздельнополые виды. (Ху и др. , 2017). Отсутствие половых партнеров может усилить зависимость от бесполого размножения, что приведет к дальнейшему сокращению разнообразия генотипов и доступных партнеров.Однако виды, производящие бесполые пропагулы, такие как луковицы или геммы, могут не испытывать сильного слипания генотипов, вызванного вегетативным ростом. Распространение бесполых отростков внутри популяции может позволить генотипам стать встречно-пальцевыми, тем самым разбивая группы генотипов, возникающие в результате вегетативного роста (Vallejo-Marín et al. , 2010). Для ауткроссинговых видов определение пространственного распределения рамет и генетов в популяции может предоставить информацию о преобладании бесполых и бесполых видов.половое размножение. Оценка полового размножения важна для лучшего прогнозирования устойчивости популяции и поддержания генотипического разнообразия и, в конечном итоге, эволюционного потенциала.
, 2015) и может ограничить половое размножение у облигатных ауткроссинговых видов, таких как самонесовместимые и раздельнополые виды. (Ху и др. , 2017). Отсутствие половых партнеров может усилить зависимость от бесполого размножения, что приведет к дальнейшему сокращению разнообразия генотипов и доступных партнеров.Однако виды, производящие бесполые пропагулы, такие как луковицы или геммы, могут не испытывать сильного слипания генотипов, вызванного вегетативным ростом. Распространение бесполых отростков внутри популяции может позволить генотипам стать встречно-пальцевыми, тем самым разбивая группы генотипов, возникающие в результате вегетативного роста (Vallejo-Marín et al. , 2010). Для ауткроссинговых видов определение пространственного распределения рамет и генетов в популяции может предоставить информацию о преобладании бесполых и бесполых видов.половое размножение. Оценка полового размножения важна для лучшего прогнозирования устойчивости популяции и поддержания генотипического разнообразия и, в конечном итоге, эволюционного потенциала.
Клональность также может ограничивать взаимодействие между соседними популяциями за счет сокращения рассредоточения. Колонизация и распространение среди популяции в основном происходит через половые отростки (Starfinger and Stöcklin, 1996; Eriksson, 1997; Vanderpoorten et al. , 2008), вероятно, потому, что половые отростки обычно меньше, чем клональные фрагменты или бесполые отростки (Tackenberg et al. ., 2003 г.; Корпелайнен и др. , 2005). Однако в водной среде возможно распространение бесполых отростков на большие расстояния, что может привести к обмену генами между популяциями (Johansson and Nilsson, 1993), что может быть единственным распространением, происходящим, если половые отростки не образуются. Взаимодействия между популяциями, в частности, распространенность расселения, можно количественно оценить, определив пространственное распределение генетов и раметов между популяциями. Если распространение между популяциями происходит через пропагулы, образующиеся половым путем, мы прогнозируем, что один и тот же генотип не будет обнаружен в разных популяциях, и между популяциями будет низкая генетическая дифференциация. Если имеет место какое-либо распространение бесполыми пропагулами, то мы ожидаем идентичных генотипов во множестве физически не связанных популяций.
Если имеет место какое-либо распространение бесполыми пропагулами, то мы ожидаем идентичных генотипов во множестве физически не связанных популяций.
Несмотря на важность близости полов друг к другу как условия для полового размножения, исследования SGS на раздельнополых видах не всегда включают пол особи (Alberto et al. , 2005; Ramaiya et al. , 2010; Korpelainen et al. , 2013; но см. Eppley et al. , 1998; Vandepitte et al., 2009 г.; Мизуки и др. , 2010; Деринг и др. , 2016). Это упущение, скорее всего, связано с тем, что определение пола особи, как правило, ограничивается наблюдением особей в момент образования гамет, обнаружением физических связей особи неизвестного пола с другой особью известного пола или сопоставлением генотипов известного пола с генотипами. неизвестного пола. Однако включение пола в качестве переменной важно, потому что клональность может стимулировать SSS в популяциях (Melampy, 1981) и привести к популяциям, состоящим только из одного пола (McLetchie et al. , 2002 г.; Кроули и др. , 2005), предотвращая половое размножение даже при наличии нескольких генотипов. Количественное определение соотношения полов вместо определения пространственного распределения полов может привести к неверным выводам о возможности полового размножения в популяции. Например, популяции могут содержать одинаковое количество самцов и самок, но представители полов могут быть сгруппированы внутри популяции, что ограничивает половое размножение из-за ограниченного распространения мужских гамет (Eppley, 2005).
, 2002 г.; Кроули и др. , 2005), предотвращая половое размножение даже при наличии нескольких генотипов. Количественное определение соотношения полов вместо определения пространственного распределения полов может привести к неверным выводам о возможности полового размножения в популяции. Например, популяции могут содержать одинаковое количество самцов и самок, но представители полов могут быть сгруппированы внутри популяции, что ограничивает половое размножение из-за ограниченного распространения мужских гамет (Eppley, 2005).
Поскольку расселение и укоренение у половых и бесполых пропагул различаются, мы можем определить важность репродуктивного способа (полового или бесполого) для устойчивости популяции или потока генов путем количественной оценки SGS. Если производство половых или бесполых пропагул различается между популяциями, которые испытывают разные переменные окружающей среды, то производство пропагул и любой SGS, который возникает в результате, может быть связан с этой переменной окружающей среды. SGS ранее был связан с переменными окружающей среды, связанными с конкретными участками, что позволяло установить связь между этой переменной и моделями распространения половых и бесполых пропагул.Эти исследования включают самосовместимые виды (Chung et al. , 2006; Arnaud-Haond et al. , 2007 a ) и ауткроссинговые виды, в том числе самонесовместимые (Raabová et al. , 2015) и раздельнополые (Dering et al. , 2016) виды, а также в зависимости от нескольких переменных окружающей среды, включая режимы возмущения (Kostrakiewicz-Gieralt, 2013), свет (Vandepitte et al. , 2010), высоту над уровнем моря (Sun et al. ). , 2001) и размер среды обитания (Kettenring et al., 2010).
SGS ранее был связан с переменными окружающей среды, связанными с конкретными участками, что позволяло установить связь между этой переменной и моделями распространения половых и бесполых пропагул.Эти исследования включают самосовместимые виды (Chung et al. , 2006; Arnaud-Haond et al. , 2007 a ) и ауткроссинговые виды, в том числе самонесовместимые (Raabová et al. , 2015) и раздельнополые (Dering et al. , 2016) виды, а также в зависимости от нескольких переменных окружающей среды, включая режимы возмущения (Kostrakiewicz-Gieralt, 2013), свет (Vandepitte et al. , 2010), высоту над уровнем моря (Sun et al. ). , 2001) и размер среды обитания (Kettenring et al., 2010).
Размер среды обитания потенциально может иметь большое влияние на динамику между популяциями. Если качество среды обитания постоянно, большой участок среды обитания связан с более высоким уровнем иммиграции по сравнению с меньшими участками среды обитания (Johnson, 2005), и большее количество иммигрантов может привести к более уникальным генотипам и потенциально большим возможностям для спаривания. И наоборот, небольшие участки среды обитания могут иметь более ограниченную иммиграцию и меньшую генетическую изменчивость (Biere et al. , 2012), и существует большая вероятность потери одного пола, что снижает или даже устраняет потенциал спаривания.Этот сниженный потенциал спаривания может привести к большей зависимости от бесполого размножения, что затем ограничивает динамику внутри популяции и влияет на долгосрочное сохранение популяции, особенно потому, что в небольших местах обитания может наблюдаться повышенный уровень эмиграции (Thomas and Hanski, 1997; Johnson, 2005) и более подвержены демографической или экологической стохастичности (Lande, 1988).
И наоборот, небольшие участки среды обитания могут иметь более ограниченную иммиграцию и меньшую генетическую изменчивость (Biere et al. , 2012), и существует большая вероятность потери одного пола, что снижает или даже устраняет потенциал спаривания.Этот сниженный потенциал спаривания может привести к большей зависимости от бесполого размножения, что затем ограничивает динамику внутри популяции и влияет на долгосрочное сохранение популяции, особенно потому, что в небольших местах обитания может наблюдаться повышенный уровень эмиграции (Thomas and Hanski, 1997; Johnson, 2005) и более подвержены демографической или экологической стохастичности (Lande, 1988).
Используя раздельнополую мохообразную Marchantia inflexa Nees et Mont., мы количественно определили SGS генотипов внутри и среди популяций в участках среды обитания разного размера (переменная окружающей среды) и впоследствии проверили на (1) сигнатуру половой принадлежности против половой.бесполое размножение на СГС; (2) пространственная ассоциация полов через раметы внутри генетов; 3) зависимость (1) и (2) от размеров участка обитания. В популяциях, если бесполое размножение является основной формой воспроизводства, мы прогнозируем, что в целом большинство особей в популяции будут состоять из нескольких копий одного и того же генотипа. Кроме того, если бесполое размножение является основной формой размножения, то мы прогнозируем значительную генетическую дифференциацию между популяциями и высокий SGS, поскольку бесполые пропагулы обычно остаются в исходной популяции.Если половое размножение является основной формой воспроизводства в популяции, то большинство особей будут уникальными генотипами. Поскольку половые отростки могут оставаться в пределах исходной популяции, а также рассеиваться между популяциями, мы ожидаем увидеть генетическое смешение среди популяций. Мы также прогнозируем, что будет поток генов, о чем свидетельствует отсутствие значительной генетической дифференциации, поэтому низкий SGS между популяциями. В качестве альтернативы могут иметь место оба репродуктивных метода и влиять на SGS и поток генов.
В популяциях, если бесполое размножение является основной формой воспроизводства, мы прогнозируем, что в целом большинство особей в популяции будут состоять из нескольких копий одного и того же генотипа. Кроме того, если бесполое размножение является основной формой размножения, то мы прогнозируем значительную генетическую дифференциацию между популяциями и высокий SGS, поскольку бесполые пропагулы обычно остаются в исходной популяции.Если половое размножение является основной формой воспроизводства в популяции, то большинство особей будут уникальными генотипами. Поскольку половые отростки могут оставаться в пределах исходной популяции, а также рассеиваться между популяциями, мы ожидаем увидеть генетическое смешение среди популяций. Мы также прогнозируем, что будет поток генов, о чем свидетельствует отсутствие значительной генетической дифференциации, поэтому низкий SGS между популяциями. В качестве альтернативы могут иметь место оба репродуктивных метода и влиять на SGS и поток генов. В этом случае мы ожидаем наблюдать как уникальные генотипы, так и генотипы с множественными клонами внутри участков среды обитания и между ними, а уровень генетической дифференциации будет указывать на преобладание потока генов.
В этом случае мы ожидаем наблюдать как уникальные генотипы, так и генотипы с множественными клонами внутри участков среды обитания и между ними, а уровень генетической дифференциации будет указывать на преобладание потока генов.
У раздельнополых видов может быть дифференциальная группировка между полами, приводящая к SSS. Поскольку самки основного вида имеют более высокую скорость вегетативного роста и более низкую скорость образования бесполых побегов, чем самцы (McLetchie and Puterbaugh, 2000; Fuselier and McLetchie, 2002), мы прогнозируем, что самки одного и того же гена будут более сгруппированы, чем самцы. того же ген.Наконец, по сравнению с популяциями в больших участках среды обитания, мы прогнозируем, что популяции в небольших участках среды обитания будут иметь низкое генетическое разнообразие, что может быть связано с меньшими ресурсами (Kareiva, 1985), более низким уровнем иммиграции (Johnson, 2005) и/или большей восприимчивостью. к демографической стохастичности (Lande, 1988). Эти силы также могут привести к более изменчивому соотношению полов в небольших популяциях по сравнению с большими популяциями из-за двух факторов: модели сниженного полового размножения или его отсутствия, порождающего новые генотипы, и зависимости от бесполого размножения для сохранения популяции.
Эти силы также могут привести к более изменчивому соотношению полов в небольших популяциях по сравнению с большими популяциями из-за двух факторов: модели сниженного полового размножения или его отсутствия, порождающего новые генотипы, и зависимости от бесполого размножения для сохранения популяции.
МАТЕРИАЛЫ И МЕТОДЫ
Исследуемый организм
Marchantia inflexa представляет собой таллоидный печеночник (тип Marchantiophyta) с отдельными мужскими и женскими растениями (двудомными). Распространение M. inflexa простирается от юга США до севера Венесуэлы (Bischler, 1984). Marchantia inflexa встречается в виде пространственно разделенных популяций вдоль ручьев на отдельных участках субстрата (валуны и коренная порода), отделенных от других участков среды обитания водой, создавая отдельные популяции, которые достаточно удалены друг от друга, чтобы предотвратить оплодотворение между особями из разных популяций (Garcia -Рамос и др. , 2007 г.; Стиха и др. , 2016). Стадия фотосинтеза гаплоидна, определение пола находится под генетическим контролем (Bischler, 1986). Из-за наличия гетерогаметных половых хромосом ожидается, что споры (половые отростки), продуцируемые диплоидным спорофитом, созревающим на женском растении, будут иметь соотношение полов 1:1 (Bischler, 1986). Оба пола могут размножаться бесполым путем как за счет бесполых побегов, так и за счет роста. Бесполые пропагулы (гемм внутри чашечки) образуются на поверхности слоевища и распространяются водой (Вrodie, 1951).Геммы могут распространяться по воздуху путем разбрызгивания воды в чаше (рассеивание внутри участка) или вниз по течению, плавая в проточной воде (расселение среди популяции) (C. Stieha, личн. наблюдения). Растения также растут горизонтально за счет дихотомического ветвления талломов, а распад более старых тканей приводит к фрагментации и образованию физиологически независимых растений. Мужские и женские растения сезонно производят различные половые структуры, которые возвышаются над растением (антеридиофоры и архегониофоры соответственно), что позволяет четко идентифицировать пол, если они присутствуют.
, 2007 г.; Стиха и др. , 2016). Стадия фотосинтеза гаплоидна, определение пола находится под генетическим контролем (Bischler, 1986). Из-за наличия гетерогаметных половых хромосом ожидается, что споры (половые отростки), продуцируемые диплоидным спорофитом, созревающим на женском растении, будут иметь соотношение полов 1:1 (Bischler, 1986). Оба пола могут размножаться бесполым путем как за счет бесполых побегов, так и за счет роста. Бесполые пропагулы (гемм внутри чашечки) образуются на поверхности слоевища и распространяются водой (Вrodie, 1951).Геммы могут распространяться по воздуху путем разбрызгивания воды в чаше (рассеивание внутри участка) или вниз по течению, плавая в проточной воде (расселение среди популяции) (C. Stieha, личн. наблюдения). Растения также растут горизонтально за счет дихотомического ветвления талломов, а распад более старых тканей приводит к фрагментации и образованию физиологически независимых растений. Мужские и женские растения сезонно производят различные половые структуры, которые возвышаются над растением (антеридиофоры и архегониофоры соответственно), что позволяет четко идентифицировать пол, если они присутствуют. Однако, когда половые структуры отсутствуют, пол визуально не различим, и для развития половых структур требуется выращивание растений в теплице или камере для выращивания. Для идентификации генетов (уникальных генотипов) и раметов (генотипических клонов или клономатов) генетов ранее были разработаны 12 полиморфных микросателлитных маркеров (Brzyski et al. , 2012).
Однако, когда половые структуры отсутствуют, пол визуально не различим, и для развития половых структур требуется выращивание растений в теплице или камере для выращивания. Для идентификации генетов (уникальных генотипов) и раметов (генотипических клонов или клономатов) генетов ранее были разработаны 12 полиморфных микросателлитных маркеров (Brzyski et al. , 2012).
Полевой отбор проб
Отбор проб был проведен в июле 2011 г. на 3-километровом участке реки Куэйр на острове Тринидад, Республика Тринидад и Тобаго.Репрезентативные образцы M. inflexa были получены в Ботаническом саду Миссури (Сент-Луис, Миссури, США, номера образцов M02 и M0) и в Национальном гербарии Республики Тринидад и Тобаго (Сент-Огастин, Тринидад, образец номер TRIN34616, DN McLetchie, коллекционер).
Мы провели количественную оценку паттернов SGS в популяциях, где оба пола встречаются одновременно, но где образцы растений изначально были неизвестного пола и могли быть определены в лаборатории. Этот подход позволяет избежать возможной систематической ошибки выборки только особей с половой структурой или только популяций с особями в половой фазе.Было отобрано пять пар участков среды обитания с M. inflexa , причем каждая пара состояла из небольшого участка (1–3 м в наибольшей длине), окруженного водой, и большого участка (>7 м в наибольшей длине), соединенного с банк потоков (дополнительный файл данных 1). Расстояние между каждой парой малых и больших пятен колебалось от 2,0 до 12,5 м (измерено рулеткой, табл. 1). Каждая пара патчей была отделена не менее чем на 20 м от другой пары патчей, при этом пары 1, 2 и 3 находились в пределах 200 м друг от друга, а пары 4 и 5 находились на расстоянии прибл.на расстоянии 1 км от других пар. Пары нумеруются в порядке от самой верхней пары (пара 1) до самой нижней пары (пара 5). Внутри пары маленькое пятно и большое пятно не обязательно располагались вверх или вниз по течению друг от друга.
Этот подход позволяет избежать возможной систематической ошибки выборки только особей с половой структурой или только популяций с особями в половой фазе.Было отобрано пять пар участков среды обитания с M. inflexa , причем каждая пара состояла из небольшого участка (1–3 м в наибольшей длине), окруженного водой, и большого участка (>7 м в наибольшей длине), соединенного с банк потоков (дополнительный файл данных 1). Расстояние между каждой парой малых и больших пятен колебалось от 2,0 до 12,5 м (измерено рулеткой, табл. 1). Каждая пара патчей была отделена не менее чем на 20 м от другой пары патчей, при этом пары 1, 2 и 3 находились в пределах 200 м друг от друга, а пары 4 и 5 находились на расстоянии прибл.на расстоянии 1 км от других пар. Пары нумеруются в порядке от самой верхней пары (пара 1) до самой нижней пары (пара 5). Внутри пары маленькое пятно и большое пятно не обязательно располагались вверх или вниз по течению друг от друга.
Размеры и расстояния между участками пяти пар малых и больших участков среды обитания, содержащих Marchantia inflexa , отобранных вдоль реки Куэр
Пара
. | Маленький (м) . | Большой (м) . | Расстояние между патчами (м) . |
|---|---|---|---|
| 1 | 2,5 | 16,0 | 12,5 |
| 2 | 3,5 | 25,0 | 5,0 |
| 3 | 2,1 | 7,0 | 11,0 |
| 4 | 2,6 | 7,0 | 6,0 |
| 5 | 2.0 | 12,0 | 2,0 |
| Пара . | Маленький (м) . | Большой (м) . | Расстояние между патчами (м) . | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 25 | 16.0 | 16.0 | 12.59 | 12.59 | 2 | 3,5 | 25,0 | 25,0 | 5.0 |
| 3 | 7.1 | 7.0 | 11.0 | 11.0 | ||||||
| 4 | 2 | 2 | 7. 0 0 | 6.0 | ||||||
| 5 | ||||||||||
| 5 | ||||||||||
| 5 | 2,0 | 12,0 | 2,0 |
Размеры и межприкладные расстояния пяти маленьких пары участков среды обитания, содержащие Marchantia inflexa , отобранные вдоль реки Куэр
| Пара . | Маленький (м) . | Большой (м) . | Расстояние между патчами (м) . | |
|---|---|---|---|---|
| 1 | 2,5 | 16,0 | 12,5 | |
| 2 | 3,5 | 25,0 | 5,0 | |
| 3 | 2,1 | 7,0 | 11,0 | |
| 4 | 2.6 | 7.0 | 7.0 | 6.0 |
| 5 | 2.0 | 12.0 | 2,0 | 2,0 |

Перед отбором проб определяли наибольшую длину разреза для каждого участка. Вдоль этого разреза была расположена случайная точка, а второй разрез (длиной 1 м и в пределах 1 м от случайной точки) был помещен так, чтобы пересечь как можно больше талломов M. inflexa , и был расположен от кромки воды внутрь патча. Единственным исключением был небольшой участок пары 5, где трансекта касалась воды с обоих концов. Половое выражение не происходило во время сбора.На каждом 2-сантиметровом интервале разреза, если присутствовало растение, мы собирали один растущий кончик слоевища, что в сумме может составлять 50 растений на участке (рис. 1). Большинство растений были маленькими (<5 мм), и ни один из разрезов не пересекал маты перекрывающихся талломов. Растения были возвращены в США и выращены в теплице Университета Кентукки для определения пола путем мониторинга экспрессии пола и получения достаточного количества ткани для выделения ДНК. Особей, у которых не развились половые структуры в теплице, помещали в ростовую камеру (14 ч при 20 °С и 10 ч при 16 °С при 24-часовом освещении, флуоресцентном свете и лампе накаливания при температуре ок.35 мкмоль м 90 580 –2 90 581 с 90 580 –1 90 581 ) для стимулирования проявления пола. Девять растений погибли до проявления пола, а четыре никогда не выражали пол. Эти лица были исключены из всех анализов.
1). Большинство растений были маленькими (<5 мм), и ни один из разрезов не пересекал маты перекрывающихся талломов. Растения были возвращены в США и выращены в теплице Университета Кентукки для определения пола путем мониторинга экспрессии пола и получения достаточного количества ткани для выделения ДНК. Особей, у которых не развились половые структуры в теплице, помещали в ростовую камеру (14 ч при 20 °С и 10 ч при 16 °С при 24-часовом освещении, флуоресцентном свете и лампе накаливания при температуре ок.35 мкмоль м 90 580 –2 90 581 с 90 580 –1 90 581 ) для стимулирования проявления пола. Девять растений погибли до проявления пола, а четыре никогда не выражали пол. Эти лица были исключены из всех анализов.
Рис. 1.
Пространственная схема растений Marchantia inflexa , собранных на десяти трансектах по 100 см на пяти парах малых и больших участков местообитаний. Растения, если они присутствовали, собирали через каждые 2 см вдоль трансекты, при этом точка на уровне 0 см соответствовала кромке воды. Каждый уникальный генотип (генет и его генотипические клоны) представлен буквенной комбинацией. Например, генотип С отличается от генотипа СС и ССС.
Каждый уникальный генотип (генет и его генотипические клоны) представлен буквенной комбинацией. Например, генотип С отличается от генотипа СС и ССС.
Рис. 1.
Пространственная схема растений Marchantia inflexa , собранных на десяти 100-сантиметровых разрезах на пяти парах малых и больших участков местообитаний. Растения, если они присутствовали, собирали через каждые 2 см вдоль трансекты, при этом точка на уровне 0 см соответствовала кромке воды. Каждый уникальный генотип (генет и его генотипические клоны) представлен буквенной комбинацией.Например, генотип С отличается от генотипа СС и ССС.
Генотипирование
ДНК экстрагировали из талломной ткани каждого образца растения с использованием модифицированного метода бромида цетилтриметиламмония (CTAB) (адаптировано из Doyle and Doyle, 1987) и амплифицировали с использованием 12 полиморфных микросателлитных праймеров, специально разработанных для M. inflexa (Brzyski et al. , 2012). Праймеры были флуоресцентно помечены во время ПЦР, и несколько локусов были объединены для анализа фрагментов.Генотипирование проводилось в Центре передовых генетических технологий Университета Кентукки на анализаторе ДНК 3730 (Applied Biosystems, Форчун-Сити, Калифорния, США) с использованием стандарта внутреннего размера ROX 400HD. Анализ фрагментов проводили с помощью Peak Scanner версии 1.0 (Applied Biosystems).
inflexa (Brzyski et al. , 2012). Праймеры были флуоресцентно помечены во время ПЦР, и несколько локусов были объединены для анализа фрагментов.Генотипирование проводилось в Центре передовых генетических технологий Университета Кентукки на анализаторе ДНК 3730 (Applied Biosystems, Форчун-Сити, Калифорния, США) с использованием стандарта внутреннего размера ROX 400HD. Анализ фрагментов проводили с помощью Peak Scanner версии 1.0 (Applied Biosystems).
Описательная генетическая статистика
Большинство растений различались по аллелям в двух и более локусах. В 31 случае одно растение отличалось от другого только по одному локусу.Из них 13 различались по длине своих фрагментов на 2 п.н., а остальные отличались до 10 п.н., что позволяет предположить, что ошибка подсчета не была причиной этих различий по одному аллелю. Поэтому для последующего анализа эти 31 растение рассматривались как уникальные генотипы. Чтобы гарантировать, что идентичные генотипы относятся к одному и тому же клону, мы рассчитали P пол , то есть вероятность, основанную на наблюдаемых частотах аллелей, что два человека могут иметь один и тот же генотип случайно из-за полового размножения (получено из теории Арно). -Хаонд и др., 2007 б ). Когда значение P пола <0,01, идентичные генотипы можно считать относящимися к одному и тому же гену (Arnaud-Haond et al. , 2007 b ).
Чтобы гарантировать, что идентичные генотипы относятся к одному и тому же клону, мы рассчитали P пол , то есть вероятность, основанную на наблюдаемых частотах аллелей, что два человека могут иметь один и тот же генотип случайно из-за полового размножения (получено из теории Арно). -Хаонд и др., 2007 б ). Когда значение P пола <0,01, идентичные генотипы можно считать относящимися к одному и тому же гену (Arnaud-Haond et al. , 2007 b ).
Описательная статистика общей генетической изменчивости была рассчитана с использованием GDA 1.1 (Lewis and Zaykin, 2001), включая аллельное богатство (количество аллелей на полиморфный локус) и количество уникальных аллелей. Поскольку ткани растений гаплоидны, значения гетерозиготности и коэффициенты инбридинга не могли быть рассчитаны.Вместо этого мы рассчитали беспристрастное измерение разнообразия Шеннона с использованием GenAlEx 6.5 (Peakall and Smouse, 2012), которая также использовалась для идентификации многолокусных генотипов путем сопоставления локусов.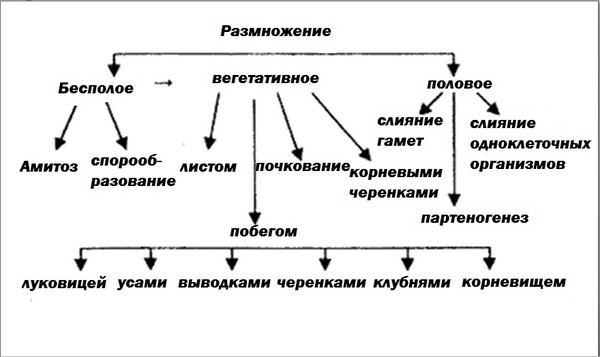
Генотипическое богатство грядки рассчитывали как G / N , где G — количество генотипов (генетов), а N — общее количество растений (раметов), отобранных по всем генотипам (Эллстранд и Руз). , 1987). Соотношение полов как для генетов, так и для рамет рассчитывали как долю самцов.Размер участка среды обитания рассматривался как категориальная переменная, обозначенная как маленькая или большая. Чтобы определить отношение размера местообитания к генотипическому богатству, аллельному богатству, уникальным аллелям и соотношению полов, мы использовали парные тесты t , в которых мы объединили маленькие и большие участки на основе нашей процедуры выборки, описанной выше. Чтобы проверить связь генотипического богатства и соотношения полов (genet и ramet) между раметами и генетами, мы использовали коэффициент корреляции Пирсона ( r ).Для анализа, включающего количество уникальных аллелей и соотношение полов, мы использовали непараметрический коэффициент Спирмена ( ρ ) из-за нарушения предположения о нормальности остатков. Наконец, общие отклонения в соотношении полов были проверены на генетическом и раметном уровнях. Чтобы проверить, было ли соотношение полов в популяциях из небольших участков более изменчивым, чем в популяциях из больших участков, мы использовали критерий Левена, чтобы определить, различаются ли различия между этими двумя группами (Vandepitte et al., 2010) на генетическом и раметном уровне.
Наконец, общие отклонения в соотношении полов были проверены на генетическом и раметном уровнях. Чтобы проверить, было ли соотношение полов в популяциях из небольших участков более изменчивым, чем в популяциях из больших участков, мы использовали критерий Левена, чтобы определить, различаются ли различия между этими двумя группами (Vandepitte et al., 2010) на генетическом и раметном уровне.
Закономерности пространственного распределения полов и генотипов в пределах участков местообитаний
Чтобы изучить пространственное распределение полов и пространственную генетическую структуру в пределах участка среды обитания, мы количественно определили, как расстояние, пол и размер участка среды обитания (маленький/большой) влияют на вероятность того, что пары особей одного пола или генотипа. Чтобы определить вероятность того, что два человека были одного пола, что соответствовало уровню SSS в пределах участка, участки должны были содержать оба пола; поэтому мы исключили из анализа пространственного распределения полов три однополых участка (маленький участок в парах пятен 2 и 5 и большой участок в паре пятен 3). Чтобы определить вероятность того, что два случайно выбранных человека были одного и того же генотипа, который измерял SGS на участке, мы использовали особей всех генотипов на всех участках. Для количественной оценки размера скопления генотипа в зависимости от пола и размера пятна мы использовали только особей, принадлежащих к генотипам с несколькими особями в одном пятне.
Чтобы определить вероятность того, что два случайно выбранных человека были одного и того же генотипа, который измерял SGS на участке, мы использовали особей всех генотипов на всех участках. Для количественной оценки размера скопления генотипа в зависимости от пола и размера пятна мы использовали только особей, принадлежащих к генотипам с несколькими особями в одном пятне.
Чтобы выполнить наш анализ и учесть SGS особей в пределах участков, мы сначала преобразовали данные разрезов в попарные сравнения между особями в пределах участков.Мы включили попарные сравнения особи с собой, т.е. когда расстояние между двумя особями равно 0 см; удаление этих сравнений качественно не повлияло на результаты. Поскольку в каждом попарном сравнении участвуют два человека, мы использовали каждое сравнение дважды при анализе наших данных. В одном случае один человек был фокусным лицом. В следующем случае другой индивидуум был фокальным индивидуумом. Для каждого попарного сравнения мы рассчитывали пространственное расстояние (в сантиметрах) между двумя людьми. Для фокальной особи мы включили ее пол, генотип, индивидуальный идентификационный номер, связанный с ее пространственным положением в пределах пятна, размер пятна и идентификационный номер пары пятен. Для каждого попарного сравнения мы определяли переменные бинарного ответа, определяющие, были ли два человека одного пола или генотипа. Поэтому для анализа наших данных мы использовали логистическую регрессию как с фиксированными, так и со случайными эффектами, используя библиотеку lme4 (Bates et al. , 2014) в R (R Core Team, 2013).Помимо анализа наших данных с упором на то, были ли два человека одного и того же генотипа, мы также проанализировали наши данные, вычислив генетическое расстояние между людьми в пределах участка, что может объяснить мутацию внутри клона и историю полового размножения (дополнительный файл данных 2; Таблица S1, рис. S1). Однако этот анализ дал результаты, аналогичные приведенному выше анализу; поэтому мы сосредоточимся на анализе того, были ли два человека одного и того же генотипа.
Для фокальной особи мы включили ее пол, генотип, индивидуальный идентификационный номер, связанный с ее пространственным положением в пределах пятна, размер пятна и идентификационный номер пары пятен. Для каждого попарного сравнения мы определяли переменные бинарного ответа, определяющие, были ли два человека одного пола или генотипа. Поэтому для анализа наших данных мы использовали логистическую регрессию как с фиксированными, так и со случайными эффектами, используя библиотеку lme4 (Bates et al. , 2014) в R (R Core Team, 2013).Помимо анализа наших данных с упором на то, были ли два человека одного и того же генотипа, мы также проанализировали наши данные, вычислив генетическое расстояние между людьми в пределах участка, что может объяснить мутацию внутри клона и историю полового размножения (дополнительный файл данных 2; Таблица S1, рис. S1). Однако этот анализ дал результаты, аналогичные приведенному выше анализу; поэтому мы сосредоточимся на анализе того, были ли два человека одного и того же генотипа.
В нашу статистическую модель мы включили фиксированные эффекты пола основной особи, размера участка среды обитания и расстояния между двумя единицами выборки, а также случайные эффекты идентификационного номера пары участков, генотипа и индивидуального идентификационного кода.Идентификационный номер пары патчей учитывал сходство между двумя патчами в паре патчей. Генотип как случайный эффект охватывал потенциальные различия жизненного цикла между генотипами. Мы включили индивидуальный идентификационный код, вложенный в генотип, чтобы учесть тот факт, что каждый отобранный индивидуум имел несколько парных сравнений. Мы использовали как случайные пересечения, так и случайные наклоны в отношении взаимосвязи между генотипом, особью и расстоянием. Различные генотипы могут иметь различия в жизненном цикле (например, более вегетативный рост или более бесполое размножение), что повлияет на то, насколько далеко особи этого генотипа могут быть рассеяны от других особей этого генотипа.Включив взаимодействие между генотипом, особью и расстоянием, мы можем сосредоточиться на поле генотипа, расстоянии между двумя особями и размере участка, который был измерен, а также их влиянии на то, принадлежит ли соседняя особь того же пола или генотипа. .
.
Чтобы определить, какие фиксированные эффекты в значительной степени объясняют вероятность того, что два человека принадлежат к одному полу или генотипу, мы провели редукцию модели (Crawley, 2007). Мы начали с полной модели, содержащей все случайные эффекты, взаимодействия между расстоянием и особью, вложенные в генотип, все фиксированные эффекты и взаимодействия между фиксированными эффектами.Чтобы определить значимость фиксированных эффектов, мы вычислили отклонение между моделью с термином и моделью без члена и проверили значимость разницы в отклонении с помощью теста х 2 . Когда условия взаимодействия были значительными, мы использовали тест z для определения различий между комбинацией фиксированных эффектов. Мы выполнили эти сравнения, установив одну комбинацию в качестве эталона и проверив, значительно ли коэффициенты для других комбинаций (и, следовательно, изменение коэффициента по сравнению с эталонным уровнем) отличались от эталона.
Модели генетической дифференциации участков среды обитания
Мы рассчитали генетическую дифференциацию между пятью парами пятен, а также между всеми пятнами с индексом взаимной информации Шеннона ( sHua ) с использованием GenAlEx 6.5 (Peakall and Smouse, 2012). Этот индекс предоставляет ту же информацию, что и F ST (Wright, 1951), но с тем преимуществом, что он более чувствителен к редким аллелям, включает размеры выборки и обеспечивает более надежную оценку дисперсии, чем другие методы (Sherwin et al., 2006 г.; Шервин, 2010). Значения генетической дифференциации варьируются от нуля до единицы, где ноль указывает на полное перекрытие генотипов во всех участках среды обитания, а единица указывает на отсутствие общих аллелей между участками. Мы также провели анализ молекулярной дисперсии (AMOVA) для количественной оценки иерархической генетической дифференциации внутри и между популяциями с использованием GenAlEx 6.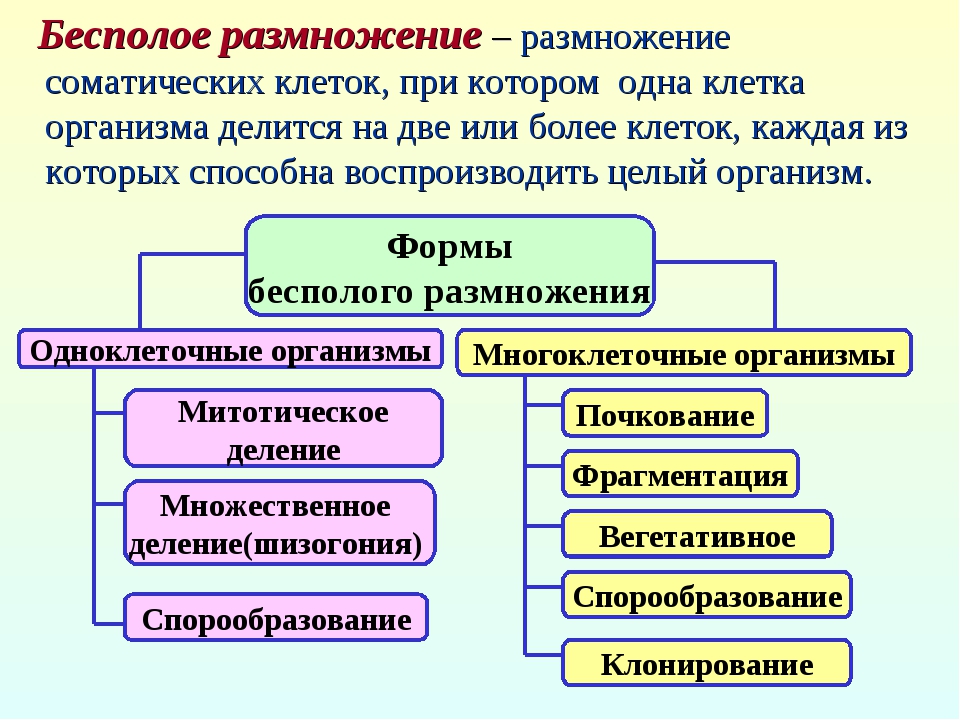 5 (Peakall and Smouse, 2012).
5 (Peakall and Smouse, 2012).
РЕЗУЛЬТАТЫ
Описательная генетическая статистика
Мы взяли образцы 128 растений с небольших участков и 98 растений с больших участков, всего 226 растений (таблица 2).После генотипирования 13 особей содержали хотя бы один локус, который не амплифицировался успешно, и были удалены из всех анализов. Из оставшихся 213 было 79 мультилокусных генотипов, из которых 34 имели несколько раметов, а 45 были одиночными уникальными генотипами (рис. 1). Вероятность образования идентичного генотипа в результате различных половых репродуктивных актов была значительно низкой ( P секс < 0,001), что позволяет сделать вывод, что особи одного и того же генотипа являются продуктами бесполого размножения.
Таблица 2.Генетическая описательная статистика Marchantia inflexa , собранная в каждой паре участков среды обитания, включая размер выборки, среднее число аллелей на локус и общее количество аллелей, уникальных для этого участка, беспристрастное измерение разнообразия Шеннона, генотипическое богатство ( G / N ) и соотношение полов (доля самцов) для потомков и рамет
Субстратная пара
. | Размер подложки . | п . | Аллели/локус . | Уникальные аллели . | Разнообразие . | Г / Н . | Генетическое соотношение полов . | Соотношение полов Рамета . | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Маленький | 24 | 2,90 | 3 | 0,336 | 33 | 0,75 | 0,92 | |||
| Большой | 38 | 2,73 | 1 | 0,343 | 0,63 | 0,67 | 0,63 | ||||
| 2 | Малый | 28 | 2,14 | 0 | 0 | 0 | 0.339 | 0,14 | 0 | 0 | 0 |
| Большой | 13 | 13 | 2,56 | 0 | 0.088 | 0,31 | 0,50 | 0,15 | |||
| 3 | Малый | 22 | 2,45 | 0 | 0,277 | 0,36 | 0,38 | 0,45 | |||
| Большой | 8 | 2,00 | 0 | 0 | 0 | 0 | 0. 330 330 | 0,25 | 1.00 | 1,00 | |
| 4 | |||||||||||
| 4 | Small | 20 | 2.33 | 3 | 0.522 | 0,25 | 0,40 | 0,20 | |||
| Большой | 16 | 3,12 | 2 | 0,260 | 0,56 | 0,78 | 0,75 | ||||
| 5 | Малый | 25 | 2.81 | 1 | 1 | 0.266 | 0.32 | 0 | 0 | ||
| Большой | 19 | 2,00 | 0 | 0.271 | 0,37 | 0,43 | 0,58 |
| Пара подложек . | Размер подложки . | п . | Аллели/локус . | Уникальные аллели . | Разнообразие . | Г / Н
. | Генетическое соотношение полов . | Соотношение полов Рамета . | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Малый | 24 | 2,90 | 3 | 0,336 | 0,33 | 0,75 | 0,92 | |||
| Большой | 38 | 2,73 | 1 | 0,343 | 0.63 | 0.67 | 0.67 | 0.63 | 0.63 | ||
| 2 | 2 | 28 | 28 | 28 | 2.14 | 0 | 0 | 0.339 | 0.14 | 0 | 0 |
| Большой | 13 | 2,56 | 0 | 0,088 | 0,31 | 0,50 | 0,15 | ||||
| 3 | Малый | 22 | 2,45 | 0 | 0.977 | 0.277 | 0.36 | 0.38 | 0.38 | 0.45 | |
| Большой | 8 | 8 | 2,00 | 0 | 0.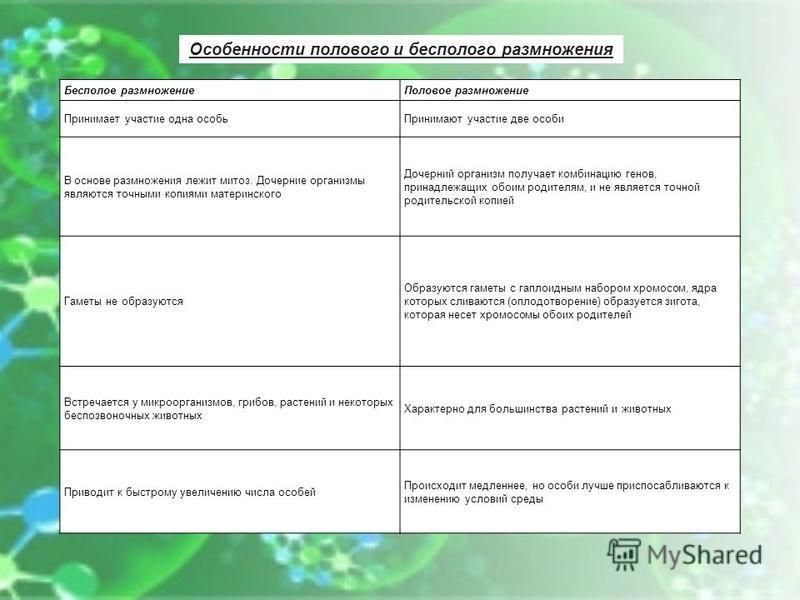 330 330 | 0,25 | 1,00 | 1,00 | |||
| 4 | Малый | 20 | 2,33 | 3 | 0,522 | 0,25 | 0,40 | 0,20 | |||
| Большой | 16 | 3.12 | 2 | 2 | 0.260 | 0.56 | 0.78 | 0.75 | 0,75 | ||
| 5 | Маленький | 25 | 2.81 | 1 | 0,266 | 0,32 | 0 | 0 | |||
| Большой | 19 | 2,00 | 0 | 0,271 | 0,37 | 0,43 | 0,58 |
Генетическая описательная статистика Marchantia inflexa , собранная в каждой паре участков среды обитания, включая размер выборки, среднее число аллелей на локус и общее количество аллелей, уникальных для этого участка, беспристрастное измерение разнообразия Шеннона, генотипическое богатство ( G / N ) и соотношения полов (доля самцов) для потомков и рамет
Субстратная пара
. | Размер подложки . | п . | Аллели/локус . | Уникальные аллели . | Разнообразие . | Г / Н . | Генетическое соотношение полов . | Соотношение полов Рамета . | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Маленький | 24 | 2,90 | 3 | 0.336 | 0,33 | 0,75 | 0,92 | |||
| Большой | 38 | 2,73 | 1 | 0,343 | 0,63 | 0,67 | 0,63 | ||||
| 2 | Малый | 28 | 2.14 | 0 | 0 | 0 | 0 939 | 0 | 0 | 0 | |
| Большой | 13 | 256 | 0 | 0.088 | 0,31 | 0,50 | 0,15 | ||||
| 3 | Малый | 22 | 2,45 | 0 | 0,277 | 0,36 | 0,38 | 0,45 | |||
| Большой | 8 | 2,00 | 0 | 0 | 0 | 0 | 0. 330 330 | 0,25 | 1.00 | 1,00 | |
| 4 | |||||||||||
| 4 | Small | 20 | 2.33 | 3 | 0.522 | 0,25 | 0,40 | 0,20 | |||
| Большой | 16 | 3,12 | 2 | 0,260 | 0,56 | 0,78 | 0,75 | ||||
| 5 | Малый | 25 | 2.81 | 1 | 1 | 0.266 | 0.32 | 0 | 0 | ||
| Большой | 19 | 2,00 | 0 | 0.271 | 0,37 | 0,43 | 0,58 |
| Пара подложек . | Размер подложки . | п . | Аллели/локус . | Уникальные аллели . | Разнообразие . | Г / Н
. | Генетическое соотношение полов . | Соотношение полов Рамета . | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Малый | 24 | 2,90 | 3 | 0,336 | 0,33 | 0,75 | 0,92 | |||
| Большой | 38 | 2,73 | 1 | 0,343 | 0.63 | 0.67 | 0.67 | 0.63 | 0.63 | ||
| 2 | 2 | 28 | 28 | 28 | 2.14 | 0 | 0 | 0.339 | 0.14 | 0 | 0 |
| Большой | 13 | 2,56 | 0 | 0,088 | 0,31 | 0,50 | 0,15 | ||||
| 3 | Малый | 22 | 2,45 | 0 | 0.977 | 0.277 | 0.36 | 0.38 | 0.38 | 0.45 | |
| Большой | 8 | 8 | 2,00 | 0 | 0. 330 330 | 0,25 | 1,00 | 1,00 | |||
| 4 | Малый | 20 | 2,33 | 3 | 0,522 | 0,25 | 0,40 | 0,20 | |||
| Большой | 16 | 3.12 | 2 | 2 | 0.260 | 0.56 | 0.78 | 0.75 | 0,75 | ||
| 5 | Маленький | 25 | 2.81 | 1 | 0,266 | 0,32 | 0 | 0 | |||
| Большой | 19 | 2,00 | 0 | 0,271 | 0,37 | 0,43 | 0,58 |
общее среднее аллельное богатство составляло 2,5 аллеля/локус на участок (диапазон 2,00–3,12; таблица 2). Небольшие участки имели в среднем более высокие числовые значения аллельного богатства (2,53 по сравнению с 2,48), уникальных аллелей (семь по сравнению с тремя) и разнообразия (0.35 по сравнению с 0,26), чем большие пятна, но ни одна из этих переменных не отличалась статистически и не коррелировала друг с другом ( ρ = 0,37–0,57, d.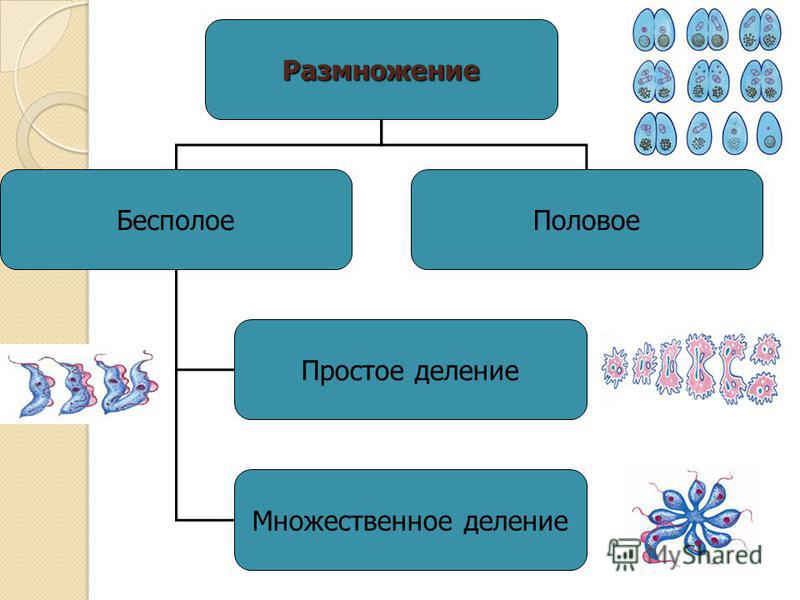 f. = 9, P > 0,2). Среднее генотипическое богатство пятен составляло 0,35 (диапазон 0,14–0,63; таблица 2). Во всех парах пятен, кроме одной, маленькие пятна имели более низкое генотипическое богатство (среднее значение = 0,28), чем большие пятна (среднее значение = 0,42), но это не было статистически значимым (парный тест t , df = 4, P = 0,14). ).
f. = 9, P > 0,2). Среднее генотипическое богатство пятен составляло 0,35 (диапазон 0,14–0,63; таблица 2). Во всех парах пятен, кроме одной, маленькие пятна имели более низкое генотипическое богатство (среднее значение = 0,28), чем большие пятна (среднее значение = 0,42), но это не было статистически значимым (парный тест t , df = 4, P = 0,14). ).
Три популяции содержали только один пол, один большой участок, состоящий только из самцов, и два небольших участка, состоящий только из самок, и все они содержали более одного генотипа (рис.1; Таблица 2). В целом, генетическое соотношение полов (0,52, доля мужчин) не отличалось от 0,5 ( х 2 = 0,1139, df = 1, P = 0,736), а раметное соотношение полов (0,44) имело тенденцию к женскому смещению (0,44). х 2 = 3,4318, df = 1, P = 0,064). Основываясь на парах участков, соотношение полов генетов было более склонным к самцам на больших участках, чем на маленьких участках (парный тест t , df = 4, P = 0,04), но соотношение полов рамет не было связано с размером участка (парный т -испытание, д. ф. = 4, P = 0,14). Дисперсия соотношения полов не отличалась между популяциями из больших и малых участков местообитаний (генетические соотношения полов F = 0,6543, df = 1, P = 0,4420; раметные соотношения полов F = 0,5251, df = 1, P = 0,4893). На всех участках соотношение полов рамет и генов сильно коррелировало друг с другом ( r = 0,9072, df = 9, P <0,001).
ф. = 4, P = 0,14). Дисперсия соотношения полов не отличалась между популяциями из больших и малых участков местообитаний (генетические соотношения полов F = 0,6543, df = 1, P = 0,4420; раметные соотношения полов F = 0,5251, df = 1, P = 0,4893). На всех участках соотношение полов рамет и генов сильно коррелировало друг с другом ( r = 0,9072, df = 9, P <0,001).
Закономерности пространственного распределения полов и генотипов в пределах участков местообитаний
Вероятность того, что два человека были одного пола, зависела от пола фокального человека, размера пятна и расстояния между двумя людьми, но не зависела от взаимодействия между этими основными эффектами (таблица 3).По мере увеличения расстояния между двумя особями вероятность того, что они были одного пола, уменьшалась с примерно 0,7 на расстоянии 0 см до примерно 0,4 на расстоянии 100 см между особями (рис. 2), что предполагает пространственное разделение полов. Однако пространственное разделение полов было асимметричным по отношению к полу и размеру участка. Вероятность того, что самцы будут находиться рядом с другими самцами, примерно на 28 % выше, чем у самок, находящихся рядом с другими самками (значительный основной эффект пола фокальной особи; таблица 3).Особи на небольших участках примерно на 46 % чаще находились рядом с особями того же пола, чем особи на больших участках (значительное основное влияние размера участка; таблица 3).
2), что предполагает пространственное разделение полов. Однако пространственное разделение полов было асимметричным по отношению к полу и размеру участка. Вероятность того, что самцы будут находиться рядом с другими самцами, примерно на 28 % выше, чем у самок, находящихся рядом с другими самками (значительный основной эффект пола фокальной особи; таблица 3).Особи на небольших участках примерно на 46 % чаще находились рядом с особями того же пола, чем особи на больших участках (значительное основное влияние размера участка; таблица 3).
Статистические тесты как для сравнения полов, так и для сравнения генотипов
| . | Срок . | Базовая модель . | Δотклонение . | д.ф. . | П . | ||
|---|---|---|---|---|---|---|---|
| Сравнение между полами | Расстояние × Секс × Размер | Расстояние + Секс + Размер + Расстояние × Секс + Расстояние × Размер + Секс × Размер | 1. 0756 0756 | 1 | 0.2997 | ||
| Пол × размер | Расстояние + пол + размер + расстояние × пол + расстояние × размер | 0,2151 | 1 | 0.6428 | |||
| Расстояние × Секс | Расстояние + Секс + Размер + Расстояние × Размер | 1 | 0.3042 | 0.3042 | 9032 | ||
| Расстояние × Размер | Расстояние + Секс + Размер | 1.6034 | 1.6034 | 1.6034 | 1|||
| Размер | Расстояние + секс | 7,4035 | 1 | 0,00651 | |||
| SSEx | Расстояние + размер | 7.6285 | 1 | 0,005745 | |||
| Расстояние | Пол + Размер | 5,0022 | 1 | 0,02531 | |||
| Сравнение между генотипами | Расстояние × пол × размер † | Расстояние + пол + размер + расстояние × пол + расстояние × размер + пол × размер | 10,735 | 1 | 0,0011 | ||
| × размер пола + расстояние размер + пол | размер 0. 8148 8148 | 1 | 0,3667 | ||||
| Расстояние × размер | Расстояние + секс + Размер + пол × размер | 15,188 | 1 | <0,0001 | |||
| Пол × размер | Расстояние + пол + размер + расстояние × размер | 5,5102 | 1 | 0,0189 |
 2151
2151  735
735  0756
0756  02531
02531 | . | Срок . | Базовая модель
. | Δотклонение . | д.ф. . | П . | ||
|---|---|---|---|---|---|---|---|
| Сравнение между полами | Расстояние × Секс × Размер | Расстояние + Секс + Размер + Расстояние × Секс + Расстояние × Размер + Секс × Размер | 1.0756 | 1 | 0.2997 | ||
| Секс × Размер | Расстояние + Секс + Размер + Расстояние × Секс + расстояние × Размер | 1 | 0.6428 | ||||
| 9032 4 | Расстояние × Пол | Расстояние + Секс + Размер + Расстояние × Размер | 1.0556 | 1 | 0,3042 | ||
| Расстояние × размер | Расстояние + секс + размер | 1,6034 | 1 | 0,2054 | |||
| Размер | Расстояние + секс | 7,4035 | 1 | | 0. | 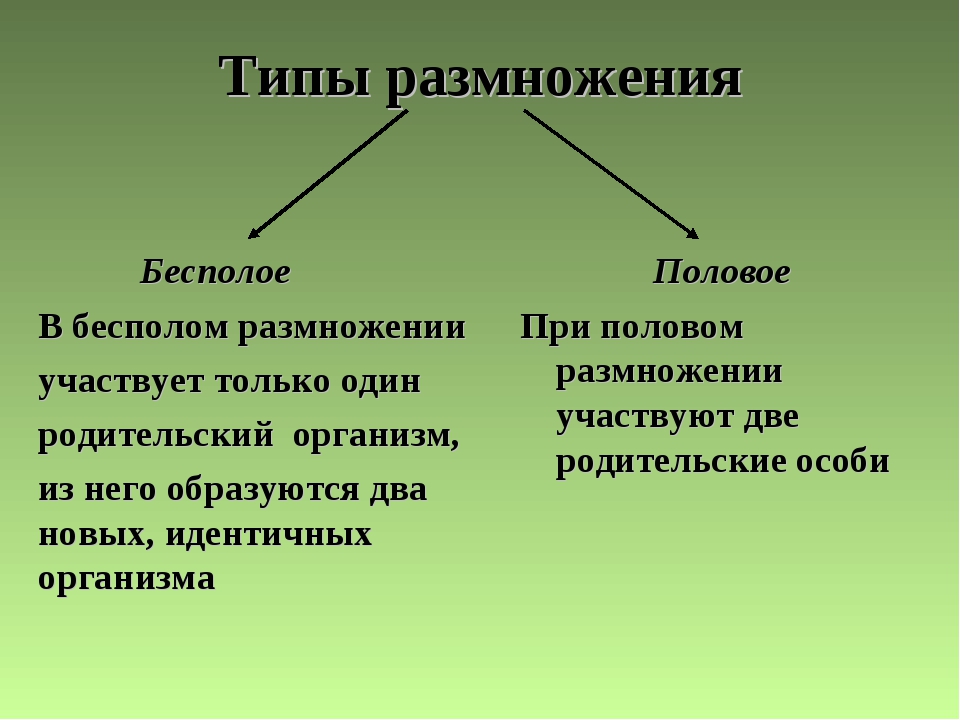 00651 00651 | ||
| SSEx | Расстояние + Размер | 7.6285 | 1 | 0.005745 | |||
| Расстояние | 60325 | 50325 | 50022 | 1 | 0,02531 | Сравнение среди генотипов | Расстояние × Секс × Размер † | Расстояние + Секс + Размер + Расстояние × Секс + Расстояние × Размер + Секс × Размер | 10.735 | 1 | 0.0011 |
| Расстояние × Секс | Расстояние + Секс + Размер + Расстояние × Размер + секс × Размер | 0.8148 | 1 | 0,3667 | |||
| Расстояние × размер | Расстояние + секс + Размер + пол × размер | 15,188 | 1 | <0,0001 | |||
| Пол × размер | Расстояние + пол + размер + расстояние × размер | 5,5102 | 1 | 0,0189 |
Рис. 2.
2.
По мере увеличения расстояния между двумя особями вероятность того, что они одного пола, уменьшалась.В наших популяциях мы наблюдали скопление полов, когда особи, находящиеся в непосредственной близости, с большей вероятностью были одного пола (пунктирная черная линия), чем ожидалось в полностью смешанной популяции (пунктирная серая линия). Мы контролировали размер участка обитания и пол фокусной особи, но эти факторы не зависели от расстояния (см. табл. 3). Черные пунктирные линии — это бутстрапированные верхний и нижний 95% доверительные интервалы, рассчитанные на основе 1000 симуляций с использованием функции bootMer в пакете lme4 (Bates et al., 2014). Прямоугольники представляют собой необработанные данные о том, были ли пары людей одного пола ( y = 1) или противоположного пола ( y = 0). Мы добавили джиттер по обеим осям, чтобы сделать все данные видимыми. Чтобы иметь возможность отобразить расстояние = 0 на графике log10, мы вычислили log10(расстояние + 1) и скорректировали метки оси x , чтобы учесть это преобразование.
Рис. 2.
По мере увеличения расстояния между двумя особями вероятность того, что они одного пола, уменьшалась.В наших популяциях мы наблюдали скопление полов, когда особи, находящиеся в непосредственной близости, с большей вероятностью были одного пола (пунктирная черная линия), чем ожидалось в полностью смешанной популяции (пунктирная серая линия). Мы контролировали размер участка обитания и пол фокусной особи, но эти факторы не зависели от расстояния (см. табл. 3). Черные пунктирные линии — это бутстрапированные верхний и нижний 95% доверительные интервалы, рассчитанные на основе 1000 симуляций с использованием функции bootMer в пакете lme4 (Bates et al., 2014). Прямоугольники представляют собой необработанные данные о том, были ли пары людей одного пола ( y = 1) или противоположного пола ( y = 0). Мы добавили джиттер по обеим осям, чтобы сделать все данные видимыми. Чтобы иметь возможность отобразить расстояние = 0 на графике log10, мы вычислили log10(расстояние + 1) и скорректировали метки оси x , чтобы учесть это преобразование.
Хотя на вероятность того, что два человека были одного пола, влияли только основные эффекты, вероятность того, что два человека были одного и того же генотипа, зависела от взаимодействия размера пятна как с расстоянием, так и с полом фокального человека (Таблица 4) .Вероятность того, что две особи имеют один и тот же генотип, уменьшалась до нуля, когда расстояние между двумя особями становилось больше 6 см; эта вероятность уменьшалась быстрее на небольших участках, чем на больших участках (рис. 3). Эффекты размера пятна также зависели от пола очага. Самцы на больших участках примерно на 13 % чаще оказывались рядом с тем же генотипом по сравнению с самцами на маленьких участках ( z = –1,88, P = 0,060) и примерно на 15 % чаще, чем самки на больших участках (). г = –2.23, P = 0,026). Самки на небольших участках не имели значительно большей или меньшей вероятности находиться рядом с особями того же генотипа, чем особи из разных комбинаций пола и участков (по сравнению с самками на больших участках, по сравнению с самцами на небольших участках и по сравнению с самцами на больших участках).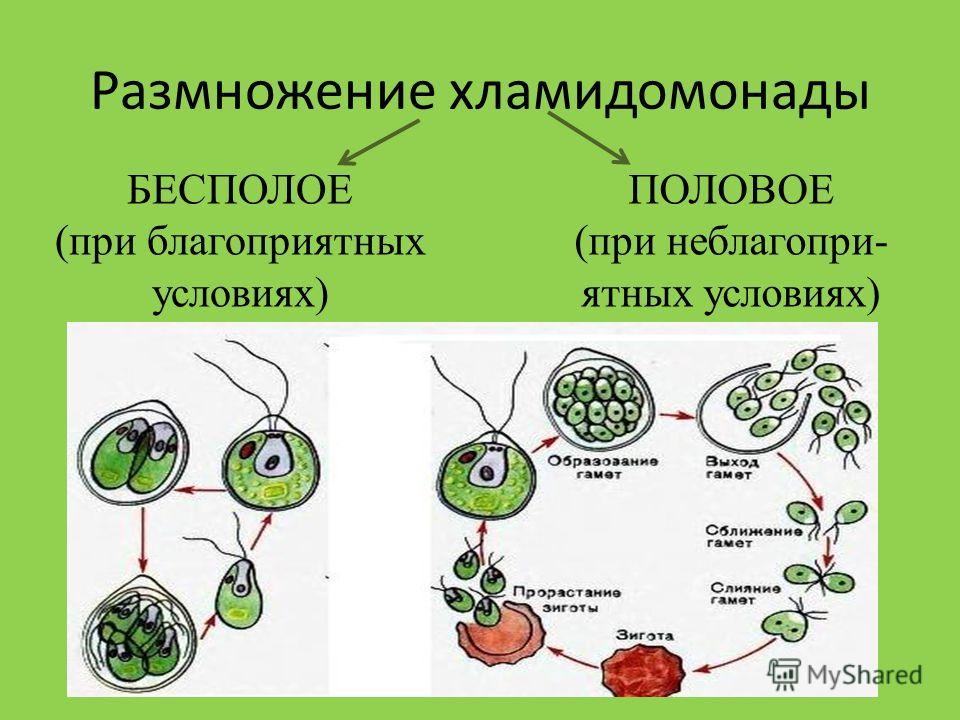 ; все P > 0,19).
; все P > 0,19).
( sHua ) популяций Marchantia inflexa
| sHua . | 1С . | 1л . | 2С . | 2л . | 3С . | 3л . | 4С . | 4л . | 5С . | 5л . | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1S | ||||||||||||
| 1л | 0227 | |||||||||||
| 2S | 0,255 | 0,193 | ||||||||||
| 2L | 0. 280 280 | 0.227 | 0,082 | |||||||||
| 3 | 0268 | 0,140 | 0,196 | 0,232 | ||||||||
| 3L | 0,441 | 0,323 | 0,421 | 0,502 | + 0,383 | |||||||
| 4S | 0,243 | 0,167 | 0,173 | 9,0327 0,173 4 0,2324В 2000,371 | ||||||||
| 4L | 0,262 | 0,234 | 0,290 | 0,316 | 0,238 | 0,570 | 0,248 | |||||
| 5s | 0.281 | 0.246 | 0.145 | 0.186 | 0.206 | 0.465 | 0. 465 465 | 0.253 | 0.293 | |||
| 5L | 0.259 | 0.132 | 0.168 | 0.232 | 0.163 | 0.459 | 0.165 | 0.348 | 0.215 |
| sHua . | 1S . | 1L . | 2S . | 2L . | 3С . | 3л . | 4С . | 4л . | 5С . | 5л . | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1S | |||||||||||||
| 1L | 0,227 | ||||||||||||
| 2S | 0. 255 255 | 0,193 | + | ||||||||||
| 2L | 0,280 | 0,227 | + 0,082 | ||||||||||
| 3S | 3S | 0.268 | 0.140 | 0.196 | 0.232 | 3l | 0441 | 0,323 | 0,421 | 0,502 | 0,383 | ||
| 4S | 0,243 | 0,167 | 0,173 | 0,257 | 0,200 | 0,371 | 9032 9 | ||||||
| 4L | 0.262 | 0.262 | 0.234 | 0.290 | 0.316 | 0238 | 0,570 | 0,248 | |||||
| 5S | 0,281 | 0,246 | 0,145 | 0,186 | 0,206 | 0,465 | 0,253 | 0,293 | |||||
| 5L | 0. 259 259 | 0.132 | 0.168 | 0.232 | 0.232 | 0.163 | 0,459 | 0,165 | 0348 | | 0.215 | | |
Генетическая дифференцировка ( Shua ) Marchantia Inflexa Население
| Shua . | 1С . | 1л . | 2С . | 2л . | 3С . | 3л . | 4С . | 4л . | 5С . | 5л . | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1S | |||||||||||||
| 1L | 0,227 | ||||||||||||
| 2S | 0. 255 255 | 0,193 | + | ||||||||||
| 2L | 0,280 | 0,227 | + 0,082 | ||||||||||
| 3S | 3S | 0.268 | 0.140 | 0.196 | 0.232 | 3l | 0441 | 0,323 | 0,421 | 0,502 | 0,383 | ||
| 4S | 0,243 | 0,167 | 0,173 | 0,257 | 0,200 | 0,371 | 9032 9 | ||||||
| 4L | 0.262 | 0.262 | 0.234 | 0.290 | 0.316 | 0238 | 0,570 | 0,248 | |||||
| 5S | 0,281 | 0,246 | 0,145 | 0,186 | 0,206 | 0,465 | 0,253 | 0,293 | |||||
| 5L | 0.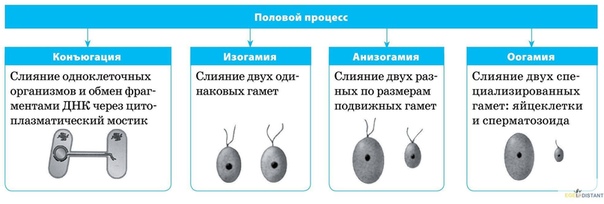 259 259 | 0.132 | 0.168 | 0.232 | 0.232 | 0.163 | 0,459 | 0,165 | 0348 | 0,215 |
| шуа . | 1С . | 1л . | 2С . | 2л . | 3С . | 3л . | 4С . | 4л . | 5С . | 5л . | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1S | ||||||||||||
| 1L | 0,227 | |||||||||||
| 2S 0,255 | 0,193 | |||||||||||
2L 0. 280 280 | 0,227 | 0.082 | ||||||||||
| 3S | 0,268 | 0,140 | 0,196 | 0,232 | ||||||||
| 3L 0,441 | 0,323 0,421 0,502 | 0,383 91 759 | ||||||||||
| 4S 0.243 | 0,167 | 0,173 | 0,257 | 0,200 | 0,371 | |||||||
| 4L | 0,262 | 0,234 | 0,290 | 0,316 | 0,238 | 0,570 | + 0.248 | |||||
| 5с | 0.281 | 0,246 | 0,145 | 0,145 | 0186 | 0,206 | 0,465 | 0,253 | 0,293 | |||
| 5л | 0,259 | 0,132 | 0,168 | 0,232 | 0,163 | 0,459 | 0,165 | 0,348 | 0,215 |
Рис. 3.
3.
Вероятность того, что две особи (раметы) имеют один и тот же генотип, уменьшалась по мере увеличения расстояния между двумя особями.На эту взаимосвязь влиял размер участка: особи на небольших участках (черная пунктирная линия) с меньшей вероятностью были почти генетически идентичными особями, чем люди на больших участках (черная пунктирная линия). Если бы особи были случайным образом распределены в пределах участка (нулевая модель), вероятность того, что любые две особи имеют один и тот же генотип, составила бы около 0,1, несмотря на расстояние между особями. Этот результат означает, что для всех особей (раметов) и генотипов (генетов) происходит скопление генотипов, и пространственная протяженность скопления генетов меньше для небольших пятен, чем для больших пятен.Поскольку многие генотипы представлены несколькими особями в пределах участка, мы можем объяснить генотипы, обнаруженные только у одной особи, и сосредоточиться на пространственном сгущении генотипа. Эти особи неравномерно распределены по всему участку (нулевая модель — серая, сплошная линия), а образуют скопления шириной до 40 см (черная, сплошная линия). Прямоугольники показывают, имеют ли два человека в каждом попарном сравнении один и тот же генотип ( y = 1) или разные генотипы ( y = 0).Мы добавили дрожание как по оси x , так и по оси y , чтобы можно было наблюдать все точки данных. Расстояние отображается как log10 (расстояние + 1), чтобы мы могли построить расстояние 0 см; метки на оси x соответствуют преобразованию.
Рис. 3.
Вероятность того, что две особи (раметы) имеют один и тот же генотип, уменьшалась по мере увеличения расстояния между двумя особями. На эту взаимосвязь влиял размер участка: особи на небольших участках (черная пунктирная линия) с меньшей вероятностью были почти генетически идентичными особями, чем люди на больших участках (черная пунктирная линия).Если бы особи были случайным образом распределены в пределах участка (нулевая модель), вероятность того, что любые две особи имеют один и тот же генотип, составила бы около 0,1, несмотря на расстояние между особями. Этот результат означает, что для всех особей (раметов) и генотипов (генетов) происходит скопление генотипов, и пространственная протяженность скопления генетов меньше для небольших пятен, чем для больших пятен. Поскольку многие генотипы представлены несколькими особями в пределах участка, мы можем объяснить генотипы, обнаруженные только у одной особи, и сосредоточиться на пространственном сгущении генотипа.Эти особи неравномерно распределены по всему участку (нулевая модель — серая, сплошная линия), а образуют скопления шириной до 40 см (черная, сплошная линия). Прямоугольники показывают, имеют ли два человека в каждом попарном сравнении один и тот же генотип ( y = 1) или разные генотипы ( y = 0). Мы добавили дрожание как по оси x , так и по оси y , чтобы можно было наблюдать все точки данных. Расстояние отображается как log10 (расстояние + 1), чтобы мы могли построить расстояние 0 см; метки на оси x соответствуют преобразованию.
Когда мы определили размер скопления генотипа, сосредоточив внимание на фокальных особях, чей генотип наблюдался более одного раза в пределах пятна, мы обнаружили, что вероятность того, что две особи имеют один и тот же генотип, достигала нуля, когда расстояние между двумя особями приближалось к 40 см (Δдевианс = 23,98, df = 1, P < 0,0001; рис. 3) по сравнению с расстоянием 6 см, когда использовались все фокальные особи. Ни участок, ни пол, ни какие-либо условия взаимодействия не были значимыми ( P > 0.16 для всех взаимодействий и основных эффектов, кроме расстояния; анализ не показан).
Закономерности генетической дифференциации участков среды обитания
Из 53 идентифицированных аллелей десять (18,9 %) были общими для всех участков среды обитания. Пять генотипов были обнаружены более чем на одном участке без какой-либо последовательной закономерности (один мужской и один женский генотип были обнаружены на обоих участках в пределах пары, в то время как три генотипа, два женских и один мужской, были обнаружены в парах участков), но не было ни одного генотипа. генотипы обнаружены более чем в двух участках.
Генетическая дифференциация между участками внутри пар участков в среднем составила 0,23, а при усреднении по всем участкам — 0,27 (диапазон = 0,08–0,57; таблица 4). Результаты AMOVA показали, что 64 % существующей генетической изменчивости было обнаружено внутри популяций и 36 % среди популяций ( P < 0,001).
ОБСУЖДЕНИЕ
В клональной раздельнополой системе Marchantia inflexa мы задокументировали как пространственное разделение полов (SSS), так и пространственное генетическое структурирование (SGS) генотипов, различающихся по полу.Среди участков обитания мы наблюдали сегрегацию как генотипов, так и полов (см. различия в соотношении полов между участками). В пределах участков среды обитания полы были пространственно разделены, при этом самцы были более сгруппированы, чем самки, тогда как генотипы также были сгруппированы, но степень сгущения зависела от взаимодействия с размером участка среды обитания и полом. Из наших результатов мы обнаружили, что среди популяций на распределение генотипов влияют как половые, так и бесполые пропагулы, в то время как на динамику внутри популяции сильно влияет бесполое размножение.
Динамика внутри популяции, особенно поток генов между участками, была генетически продемонстрирована как минимальная. Наши данные предполагают ограниченный поток генов, о чем свидетельствует наличие значительной генетической изменчивости среди пятен (36%) и относительно высокий уровень генетической дифференциации ( sHua ) среди пятен (в среднем = 0,27 ± 0,02). Большие участки среды обитания, как правило, имеют более высокое генотипическое разнообразие, чем небольшие участки, что позволяет предположить, что половое размножение чаще происходит на больших участках или что большие участки принимают больше иммигрантов, чем маленькие участки.Поскольку пространственное расстояние между генами короче на небольших участках, что предполагает лучший потенциал полового размножения на меньших, чем на больших участках, мы подозреваем, что иммиграция является наиболее вероятным объяснением этой тенденции. Обычно считается, что бесполые пропагулы не способствуют распространению на большие расстояния и более важны для внутрипопуляционной динамики семенных растений (Starfinger and Stöcklin, 1996; Santamaría, 2002). Как сообщается для других мохообразных (Laaka-Lindberg et al. , 2003; Pohjamo et al., 2006), наше исследование показало, что эти пропагулы действительно могут влиять на динамику внутри популяции, распространяясь между популяциями через воду, о чем свидетельствуют идентичные генотипы, наблюдаемые в популяциях. Мы обнаружили пять случаев бесполого расселения, происходящих либо между парами участков 1, 2 и 3, либо между парами участков 4 и 5. Все участки 1–3 были расположены в пределах 200-метрового участка реки и были отделены от участков 4 и 5 примерно . 1 км. Эти расстояния предполагают потенциально большое расстояние распространения бесполых пропагул до 200 м, но <1 км.Кривые разрежения (данные не показаны) показали, что наш протокол выборки был адекватным для количественной оценки генетической изменчивости, но, поскольку была отобрана только часть популяции, возможны дополнительные идентичные генотипы между популяциями. Имеются минимальные данные о фактическом расстоянии, которое проходят бесполые пропагулы мохообразных (Korpelainen et al. , 2005; но см. Pohjamo et al. , 2006), но моделирование показало, что нижний предел среднего путешествие бесполых побегов в м.inflexa , как ожидается, будет 2–5 м (Stieha et al. , 2014). Когда поток генов измеряется у других мохообразных (Korpelainen et al. , 2005; Paasch et al. , 2015), клональность, по-видимому, не ограничивает потенциал расселения, пока среда обитания является водной. Однако движение бесполых пропагул в нашем эксперименте, вероятно, не является обычным явлением, учитывая значительную генетическую дифференциацию популяций. По сравнению с другими значениями генетической дифференциации печеночников, которые, как сообщается, достигают 0.928 (среднее значение = 0,614 в Korpelainen et al. , 2005; 0,480 в Bączkiewicz, 2013), наше значение намного ниже, но важно отметить, что эти исследования в основном проводились в больших масштабах на сотни километров.
В популяциях преобладало бесполое размножение, которое оказывало сильное влияние на SGS. Генотипическое разнообразие, хотя и изменчивое, в среднем было низким, что свидетельствует о преобладании бесполого размножения. Как клональный рост, так и бесполые пропагулы, которые падают рядом с родительским растением, могут привести к скоплению генотипа, наблюдаемому в этом исследовании, когда особи, вероятно, будут взаимодействовать с другими особями того же генотипа на расстоянии до 6 см.Одно только это количество комков снижает возможность полового размножения (Wyatt, 1977; Reynolds, 1980; McLetchie, 1996). Однако группировка также наблюдалась у полов, вызывая пространственную сегрегацию самцов и самок и еще больше уменьшая близость к партнерам. Интересно, что SSS различались по полу и размеру участка обитания. На более крупных участках, на которых преобладали самцы, мужские генотипы были более сгущенными, чем женские генотипы, тогда как в популяциях на небольших участках среды обитания мужские и женские генотипы не различались в отношении сгущения.Эта разница в скоплении генотипов между полами указывает на влияние условий окружающей среды (включая возраст участка) на динамику нашего исследуемого организма. Выявление этих конкретных условий окружающей среды потребует дальнейших исследований.
Было показано, что воздействие окружающей среды вызывает агрегацию полов у многих видов семенных растений. Например, у двудомного многолетника Mercurialis perennis мужские гены росли больше в условиях высокой освещенности, чем женские, что приводило к сильной сегрегации полов в областях с высокой освещенностью из-за крупных мужских генов, но не сегрегации полов в условиях низкой освещенности. области (Вандепитте и др., 2010). Наши результаты демонстрируют половые различия в зависимости от размера местообитания, и это хорошо видно при сравнении средних параметров популяционного уровня больших и малых участков (табл. 2). В среднем соотношение полов, ориентированное на самцов, наблюдается на больших участках среды обитания по сравнению с небольшими участками среды обитания (0,622 ± 0,139 против 0,314 ± 0,176, среднее значение ± стандартное отклонение соответственно) и связано с большим количеством мужских генотипов, а не с большим количеством раметов на генет у самцов. В среднем большие участки местообитаний содержали в два раза больше мужских генотипов, чем женских (6.01 ± 2,67 против 3,18 ± 1,34), но самки имели преимущество по общему количеству раметов на генет (всего у самцов было 2,36 ± 0,61, а у самок 2,82 ± 0,89 раметов на генет). С другой стороны, небольшие участки местообитаний содержали в среднем вдвое меньше генотипов самцов, чем генотипов самок (2,19 ± 1,10 против 4,36 ± 1,03), а самки имели преимущество по общему количеству рамет на генет (всего самцы имели 3,00 ± 0,51 и у самок было 3,81 ± 1,09 рамет на генетику). Кроме того, две однополые популяции на небольших участках — это женщины, а одна однополая популяция на большом участке среды обитания — мужчины.Предполагая одинаковые скорости колонизации спор полами, эти закономерности предполагают более высокие показатели приживаемости или выживаемости мужских генотипов по сравнению с самками на больших участках среды обитания по сравнению с небольшими участками среды обитания, а выжившие самки, как правило, имеют больше раметов на генет, чем самцы. У семенных растений соотношение полов, ориентированное на мужчин, было связано с более высокими затратами на половое размножение у самок, где по мере старения популяции соотношение полов становится более склонным к мужчинам из-за более низкой выживаемости самок (Geber, 1999; Obeso, 2002; Field и др.). др., 2013). У M. inflexa более высокие затраты самок на половое размножение предполагают, что сперматозоиды не являются ограничивающими, но на самом деле это может быть связано с тем, что расстояние распространения сперматозоидов у мохообразных составляет порядка сантиметров (Wyatt, 1977; Reynolds, 1980; McLetchie, 1996). и мы знаем, что на этих малых расстояниях особи с большей вероятностью будут находиться рядом с особями того же пола, чем с особями противоположного пола, что приводит к неоплодотворению SSS и самок. Было высказано предположение, что ограничение количества сперматозоидов в сочетании с более высокой стоимостью предварительного оплодотворения у самцов по сравнению с самками способствует склонному к самкам соотношению полов среди мохообразных (McLetchie, 1992; Stark et al., 2000).
Мы предполагаем, что большое количество женских генетов на небольших участках среды обитания может быть результатом высокой скорости укоренения либо спорами (половые отростки), либо геммами (бесполые отростки). Самки и самцы, вероятно, имеют одинаковую скорость колонизации через половые пропагулы из-за соотношения полов спор 1: 1. Однако из-за низкого образования гемм у самок по сравнению с самцами (McLetchie and Puterbaugh, 2000), самки менее склонны к колонизации через геммы, чем самцы.Таким образом, наличие соотношения полов в популяциях на небольших участках среды обитания, ориентированное на самок, может лучше отражать более высокий уровень укоренения и устойчивости самок, а не темпы колонизации. По сравнению с самцами более высокая выживаемость самок может быть результатом более высокой устойчивости самок к обезвоживанию на стадиях геммы (Stieha et al. , 2014) и слоевища (Marks et al. , 2016). Более высокая устойчивость к обезвоживанию у самок в сочетании с большей изменчивостью условий влажности у мелких по сравнению с младшими.большие участки среды обитания, которые могут возникать в более засушливые периоды года, будут отдавать предпочтение самкам, а не самцам. Однако вариации влажности в разных размерах местообитаний еще не задокументированы.
Из-за самоподдерживающегося цикла сокращения количества партнеров посредством клонального роста бесполое размножение часто становится доминирующей формой размножения у видов со смешанным репродуктивным способом (половым и бесполым) (например, Brzyski and Culley, 2011), тем самым оказывая потенциально большое влияние как внутри, так и между патчами динамики.У раздельнополых видов пространственная сегрегация полов может значительно увеличить расстояние до ближайшего потенциального партнера, потому что даже если соседняя особь может иметь другой генотип, она может быть того же пола. Поэтому необходимо обязательно определять как пол, так и генотип особей. При этом мы смогли показать, что SSS происходит как внутри популяций, что зависит от пола и размера участка обитания, так и среди популяций, поскольку некоторые популяции имеют только один пол, а наблюдаемая нами картина соотношения полов согласуется с половыми различиями в физиология.Наконец, модель соотношения полов подтверждает интуитивное предсказание положительной связи между соотношением полов раметы и генетики, сделанное для клональных семенных растений (Eppley et al. , 1998; Vandepitte et al. , 2009; Petzold et al. ). , 2013; и см. McLetchie and García-Ramos, 2017, где представлена явная модель этой взаимосвязи у клональных двудомных растений, включающая нелинейный компонент). Количественная оценка распределения полов и распределения генотипов как внутри популяций, так и между ними дает более полную картину индивидуальной и популяционной динамики.
ДОПОЛНИТЕЛЬНЫЕ ДАННЫЕ
Дополнительные данные доступны в Интернете по адресу https://academic.oup.com/aob и включают следующее. Файл 1: схематическая диаграмма, представляющая карту пар популяций и их положение относительно друг друга и берега ручья. Файл 2: Методы и результаты вычисления генетического расстояния между особями в пределах участка. Таблица S1: анализ отношения правдоподобия. Рисунок S1: влияние пространственного расстояния на генетическое расстояние между двумя людьми.
БЛАГОДАРНОСТИ
Авторы выражают благодарность Отделу дикой природы Департамента лесного хозяйства Республики Тринидад и Тобаго за разрешения на сбор и экспорт, а также Управление водоснабжения и канализации за доступ к реке Куэр. Мы также благодарим Департамент растениеводства и почвоведения Университета Кентукки за теплицу. Финансирование было предоставлено исследовательской стипендией Университета Кентукки, присужденной Дж. Р. Б.
ЦИТИРОВАННАЯ ЛИТЕРАТУРА
ALBERTO
F
,
F
,GUVEIA
L
,
L
,S
,Pérez-Lloréns
,JL
,
JL
,Duarte
см
,Serrao
EA
.2005
.Пространственная генетическая структура внутри популяции, размер соседей и клональный подареал морских водорослей Cymodocea nodosa
.Молекулярная экология
14
:2669
–2681
.
Arnaud-Haond
S
,Migliaccio
M
,Diaz-Almela
E
, et al.2007
и .Характер викариантности в Средиземном море: расщепление с востока на запад и низкое расселение эндемичной морской травы Posidonia oceanica
.Журнал биогеографии
34
:963
–976
.
Arnaud-Haond
S
,Duarte
CM
,Alberto
F
,Serrão
4 EA 9.
2007
б .Стандартизация методов изучения клональности в популяционных исследованиях
.Молекулярная экология
16
:5115
–5139
.
Барретт
СЧ
.2015
.Влияние клональности на половое размножение растений
.Proceedings of the National Academy of Sciences, USA
112
:8859
–8866
.
Бончкевич
А
.2013
.Генетическое разнообразие видов лиственных печеночников (Jungermanniidae, Marchantiophyta) в Польше: разнообразие видов лиственных печеночников с различными способами размножения
.Биоразнообразие: исследования и сохранение
27
:3
–54
.
Бир
А
,Ван Андел
Дж
,Ван де Коппель
Дж
.2012
.Популяции: экология и генетика
. В:ван Андел
J
,Аронсон
J
, ред.Восстановление экологии: новые рубежи
, 2-е изд.Чичестер, Великобритания
:Blackwell Publishing
,73
–86
.
Бишлер
Н
.1984
.Marchantia L. Виды Нового Света
.Briophytorum Bibliotheca
26
:1
–228
.
Бишлер
Н
.1986
.Marchantia polymorpha L.s. лат. анализ кариотипа
.Журнал ботанической лаборатории Хаттори
60
:105
–117
.
Броди
ХДЖ
.1951
.Чашечный механизм в установках
.Canadian Journal of Botany
29
:224
–234
.
Бжиски
JR
,Калли
ТМ
.2011
.Генетическая изменчивость и клональная структура редкого кустарника Spiraea virginiana (Rosaceae)
.Генетика сохранения
12
:1323
–1332
.
Brzyski
JR
,ADAMS
KJ
,Walter
см
,Gale
Kh
,McLethethie
DN
.2012
.Характеристика 12 полиморфных микросателлитных маркеров в печеночнике Marchantia inflexa (Marchantiaceae)
.American Journal of Botany Prime Notes and Protocols
99
:e440
–e442
.
CHUNG
JM
,Lee
BC
,KIM
JS
,Park
CW
,Chung
My
,Chung
MG
.2006
.Мелкомасштабная генетическая структура генетических особей клонообразующего монотипического рода Echinosophora koreensis (Fabaceae)
.Анналы ботаники
98
:165
–173
.
Кроули
МДж
.2007
.Книга R
.Чичестер, Великобритания
:John Wiley and Sons
.
Crowley
PH
,Stieha
CR
,McLetchie
DN
.2005
.Конкуренция чрезмерного роста, фрагментация и динамика соотношения полов: пространственно эксплицитная субиндивидуальная модель
.Журнал теоретической биологии
233
:25
–42
. .2016
.Половой характер пространственной генетической структуры двудомных и клоновых видов деревьев, Populus alba L
.Генетика деревьев и геномы
12
:70
.
Дойл
Дж.Дж.
,Дойл
Дж.Л.
.1987
.Процедура быстрого выделения ДНК из небольшого количества тканей свежих листьев
.Фитохимический бюллетень
19
:11
–15
.
Эккерт
CG
.2000
.Вклад автогамии и гейтоногамии в самоопыление у массово цветущего клонального растения
.Экология
81
:532
–542
.
Эллстранд
NC
,Руз
ML
.1987
.Закономерности генотипического разнообразия клональных видов растений
.Американский журнал ботаники
74
:123
–131
.
Эппли
СМ
.2005
.Пространственная сегрегация полов и питательные вещества влияют на успех размножения двудомной травы, опыляемой ветром
.Экология растений
181
:179
–190
.
Эппли
СМ
,Стэнтон
МЛ
,Гросберг
РК
.1998
.Внутрипопуляционная изменчивость соотношения полов у солянки Distichlis spicata
.Американский натуралист
152
:659
–670
.
Эрикссон
О
.1997
.Истории жизни клонов и эволюция пополнения семян
.In:de Kroon
H
,van Groenendael
J
, ред.Экология и эволюция клоновых растений
.Лейден, Нидерланды
:Backhuys Publishers
,211
–226
.
Полевой
DL
,Пикап
M
,Барретт
SCH
.2013
.Сравнительный анализ изменчивости соотношения полов у раздельнополых цветковых растений
.Эволюция
67
:661
–672
.
Франкхэм
Р
.1995
.Генетика сохранения
.Annual Review of Genetics
29
:305
–327
.
Fuselier
L
,McLetchie
DN
.2002
.Сохранение полового диморфизма предвзрослых признаков у Marchantia inflexa (Marchantiacae)
.Американский журнал ботаники
89
:592
–601
.
Garcia-Ramos
G
,Stieha
C
,McLetchie
DN
,Crowley
PH
.
2007
.Сохранение полов в метапопуляциях в условиях интенсивной асимметричной конкуренции
.Журнал экологии
95
:937
–950
.
Гебер
МА
.1999
.Теории эволюции полового диморфизма
. В:Geber
MA
,Dawson
TE
,Delph
LF
, ред.Гендерный и половой диморфизм у цветковых растений
.Берлин
:Springer
,97
–122
.
Hu
A
,A
,Gale
SW
,KUMAR
P
,SAUNDERS
RMK
,SUN
M
,Fischer
GA
.2017
.Преобладание клональности вызывает потерю пола у Bulbophyllum bicolor , облигатного ауткроссинга эпифитной орхидеи
.Молекулярная экология
26
:3358
–3372
.
Йоханссон
ME
,Нильссон
С
.1993
.Гидрохория, динамика численности и распространение клонового водного растения Ranunculus lingua
.Журнал экологии
81
:81
–91
.
Джонсон
Немецкая марка
.2005
.Модели метапопуляции: эмпирическая проверка модельных допущений и методов оценки
.Экология
86
:3088
–3098
.
Карейва
Р
.1985
.Обнаружение и потеря растений-хозяев с помощью Phyllotreta : размер участка и окружающая среда обитания
.Экология
66
:1809
–1816
.
Кеттенринг
КМ
,Маккормик
МК
,Барон
ХМ
,Уигэм
DF
3.
2010
.Phragmites australis (тростник обыкновенный) инвазия в субэстуарий реки Род в Чесапикском заливе: изучение влияния лиственных питательных веществ, генетического разнообразия, размера участка и жизнеспособности семян
.Эстуарии и побережья
33
:118
–126
.
Корпелайнен
H
,Похьямо
M
,Лаака-Линдберг
S
.2005
.Насколько эффективно распространение мохообразных приводит к потоку генов
?Журнал ботанической лаборатории Хаттори
97
:195
–205
.
Корпелайнен
H
,фон Кройтляйн
M
,Костамо
K
,Виртанен
3 В 9.
2013
.Пространственная генетическая структура водных мохообразных в сообщающейся озерной системе
.Биология растений
15
:514
–521
.
Костракевич-Гиралт
К
.2013
.Влияние градиента возмущения на пополнение клональных видов растений на Molinietum caeruleae лугах
.Польский экологический журнал
61
:519
–533
.
Лаака-Линдберг
S
,Корпелайнен
H
,Похьямо
M
.2003
.Распространение бесполых побегов мохообразных
.Журнал ботанической лаборатории Хаттори
93
:319
–330
.
Lande
R
.1988
.Генетика и демография в биологической охране
.Наука
241
:1455
–1460
.
Marks
RA
,Burton
JF
,McLetchie
DN
.2016
.Половые различия и пластичность устойчивости к обезвоживанию: взгляд на тропический печеночник
.Анналы ботаники
118
:347
–356
.
Маклетчи
ДН
.1992
.Соотношение полов от прорастания до созревания и его репродуктивные последствия у печеночника Sphaerocarpos texanus
.Экология
92
:273
–278
.
Маклетчи
ДН
.1996
.Ограничение сперматозоидов и генетические эффекты на плодовитость двудомной печеночницы Sphaerocarpos texanus
.Половое размножение растений
9
:87
–92
.
McLetchie
DN
,Гарсия-Рамос
G
.2017
.Прогностическая взаимосвязь между популяцией и генетическим соотношением полов у клональных видов
.Acta Oecologia
80
:18
–23
.
Маклетчи
ДН
,Путербо
МН
.2000
.Соотношение полов в популяции, половые клональные признаки и компромиссы между этими признаками у печеночника Marchantia inflexa
.Ойкос
90
:227
–237
.
Маклетчи
DN
,Гарсия-Рамос
G
,Кроули
PH
.2002
.Локальная динамика соотношения полов: модель двудомной печеночницы Marchantia inflexa
.Эволюционная экология
15
:231
–254
.
Мелампи
МН
.1981
.Сцепленная с полом дифференциация ниш у двух видов Thalictrum
.American Midland Naturalist
106
:325
–334
.
Мизуки
I
,Исида
K
,Тани
N
,Цумура
Y
.2010
.Мелкомасштабная пространственная структура генов и полов у двудомного растения Dioscorea japonica , которое распространяется как луковицами, так и семенами
.Эволюционная экология
24
:1399
–1415
.
Обесо
JR
.2002
.Затраты на воспроизводство растений
.Новый фитолог
155
:321
–348
.
Paasch
AE
,BDchler
BD
,Nosratinia
S
,STARK
LR
,Fisher
K
.2015
.Разъединение полового размножения и генетического разнообразия у мха пустыни Мохаве, ориентированного на самок Syntrichia caninervis (Pottiaceae)
.International Journal of Plant Science
176
:751
–761
.
Peakall
R
,Smouse
PE
.2012
.GenAlEx 6.5: генетический анализ в Excel. Популяционно-генетическое программное обеспечение для обучения и исследований – обновление
.Биоинформатика
28
:2537
–2539
.
Petzold
A
,PFEiffer
,T
,T
,Jansen
F
,EuseMann
P
,Schnittler
M
.2013
.Соотношение полов и клональный рост двудомных Populus euphratica Oliv., провинция Синьцзян, Западный Китай
.Деревья
27
:729
–744
.
Похьямо
M
,Лаака-Линдберг
S
,Оваскайнен
O
,Корпелайнен
3 H 9.
2006
.Возможность распространения спор и бесполых гемм в эпиксильной печени Anastrophyllum hellerianum
.Эволюционная экология
20
:415
–430
.
Раабова
J
,Ван Россум
F
,Жакмар
AL
,Распе
O.
2015
.Размер популяции влияет на генетическое разнообразие и мелкомасштабную пространственную генетическую структуру клонального двулистного растения Menyanthes trifoliata
.Перспективы экологии, эволюции и систематики растений
17
:193
–200
.
Ramaiya
M
,Johnson
MG
,Shaw
B
, et al.2010
.Морфологически загадочные биологические виды печеночника Frullania asagrayana
.Американский журнал ботаники
97
:1707
–1718
.
R Основная группа
.2013
.R: язык и среда для статистических вычислений
.Вена, Австрия
:R Фонд статистических вычислений
.
Рейнольдс
Ду
.1980
.Распространение гамет в Mnium ciliare
.Бриолог
83
:73
–77
.
Сантамария
L
.2002
.Почему большинство водных растений широко распространены? Распространение, клональный рост и мелкомасштабная гетерогенность в стрессовой среде
.Acta oecologica
23
:137
–154
.
Шервин
ВБ
.2010
.Энтропийно-информационные подходы к генетическому разнообразию и его выражению: геномная география
.Энтропия
12
:1765
–1798
.
Шервин
ВБ
,Жабо
Ф
,Раш
Р
,Россетто
М
.2006
.Измерение биологической информации с применением от генов до ландшафтов
.Молекулярная экология
15
:2857
–2869
.
Starfinger
U
,Stöcklin
J
.1996
.Распространение семян, пыльцы и клонов и их роль в формировании популяций растений
.Прогресс в ботанике
57
:337
–355
.
Старк
ЛР
,Мишлер
БД
,Маклетчи
ДН
.2000
.Стоимость реализованного полового размножения: оценка закономерностей репродуктивного выделения и аборта спорофита у пустынного мха
.Американский журнал ботаники
87
:1599
–1608
.
Stieha
C
,García-Ramos
G
,McLetchie
DN
,Crowley
3 P 90.
2016
.Сохранение полов и персистенция клонального организма в пространственно сложных метапопуляциях
.Эволюционная экология
31
:363
–386
.
STIEHA
CR
,MIDDLELON
AR
,AR
,STIEHA
JK
,TROTT
SH
,McLethethie
DN
.2014
.Процесс распространения бесполых пропагул и вклад в сохранение популяции Marchantia (Marchantiaceae)
.Американский журнал ботаники
101
:348
–356
.
Вс
Ю
,Гао
X
,Кай
Y
.2001
.Вариации полового и бесполого размножения Scirpus mariqueteer вдоль градиента высоты
.Экологические исследования
16
:263
–274
.
Таккенберг
О
,Пошлод
P
,Бонн
S
.2003
.Оценка потенциала рассеивания ветром видов растений
.Экологические монографии
73
:191
–205
.
Томас
CD
,Хански
I
.1997
.Метапопуляции бабочек
. В:Hanski
I
,Gilpin
ME
, ред.Метапопуляционная биология
.Сан-Диего
:Academic Press
,359
–386
.
Вальехо-Марин
M
,Dorken
ME
,Barrett
SCH
.2010
.Экологические и эволюционные последствия клонирования для спаривания растений
.Ежегодный обзор экологии, эволюции и систематики
41
:193
–213
.
VANDEPITTE
K
,K
,Roldán-Ruiz
I
,LEUS
L
,Jacqueyn
H
,HONNAY
O
.2009
.Смыкание кроны формирует клональное разнообразие и мелкомасштабную генетическую структуру двудомного многолетнего подлеска Mercurialis perennis
.Журнал экологии
97
:404
–414
.
VANDEPITTE
K
,K
,,
K
,HONNAY
O
,De Meyer
T
,Jacqueyn
H
,Roldán-Ruiz
I
.2010
.Закономерности изменения соотношения полов и генетического разнообразия двудомного лесного многолетника Mercurialis perennis
.Экология растений
206
:105
–114
.
VANDERPOORTEN
A
,A
,DEVOS
N
,GOFFINET
B
,HARDY
OJ
,SHAW
AJ
.2008
.Барьеры для радиации океанических островов у мохообразных: выводы из филогеографии мхов Grimmia montana
.Журнал биогеографии
35
:654
–663
.
Райт
S
.1951
.Генетическая структура популяций
.Анналы евгеники
15
:323
–354
.
Вятт
Р
.1977
.Пространственная структура и расстояния распространения гамет у Atrichum angustatum
.Бриолог
80
:284
–291
.Примечания автора
© Автор(ы), 2018 г. Опубликовано издательством Oxford University Press от имени компании Annals of Botany. Все права защищены. Для разрешений, пожалуйста, по электронной почте: [email protected].
Объяснение важности репродукции
ФОН:Размножение важно для выживания всего живого. Без механизма воспроизводства жизнь пришла бы к конец.В начальных классах есть два типа воспроизведения: бесполое и половое размножение. Многие учителя боятся обсуждать воспроизведение, но если оно представлено в виде фактического урока, учащимся будет легко понять принципы без стандартного хихиканья.
Бесполое размножение относится к простым клеткам деление, при котором образуется точная копия организма. Есть много различные типы бесполого размножения, которые можно обсудить, чтобы показать студентов вариативность способов воспроизведения.Некоторые одноклеточные организмы размножаются простым делением клеток, это называется бинарным делением. В Таким образом, материнская клетка просто делится пополам, образуя две дочерние клетки. Некоторые клетки размножаются неравным делением клеток, это называется бутонизация. В этом процессе почка формируется как выступ на материнской клетке. То ядро делится и идентичные части идут к материнской клетке и почке. Бутон может расти до тех пор, пока не станет таким же большим, как материнская клетка, и сможет образовывать собственные почки. В некоторых случаях почки остаются прикрепленными к материнской клетке.В других они ломаются прочь и жить как отдельные организмы.
Половое размножение включает соединение мужские и женские половые клетки. Сперматозоид относится к мужской половой клетке и яйцеклетке относится к женской половой клетке.
ПРОЦЕДУРА:- Обсуждая это со студентами, скажите
им, что деревья, рыбы, цветы и многие другие организмы имеют сперму и яйцеклетки.
Предложите учащимся посмотреть на организмы по пути домой, чтобы увидеть, смогут ли они
предсказать, какие из них размножаются половым или бесполым путем.
- Есть некоторые репродуктивные стратегии
которые трудно классифицировать для учащихся. Папоротники, грибы и некоторые простейшие
размножаются спорообразованием. Споры образуются делениями специальных
материнские клетки и высвобождаются из родительского организма. Одна спора
развивается в новый организм.
- Рабочий лист показывает учащимся Отличие бесполого и полового размножения. Ответы такие следует: 1. да; 2. пополам; 3.маленькие одноклеточные существа; 4. зигота 5. женщина, мужчина, ребенок; 6. более крупные организмы с органами; 7. более крупные организмы не может просто делиться, для маленьких организмов более эффективно размножаться таким образом.
Процесс размножения в организмах на JSTOR
Перейти к основному содержанию Есть доступ к библиотеке? Войдите через свою библиотекуВесь контент Картинки
Поиск JSTOR Регистрация Вход-
Поиск
- Расширенный поиск
- Изображения
-
Просматривать
-
По тематике
Журналы и книги -
По названию
Журналы и книги - Издатели
- Коллекции
- Изображения
-
По тематике
-
Инструменты
- Рабочее пространство
- Анализатор текста
- Серия JSTOR Understanding
- Данные для исследований
Бесполое размножение у голотурий
Обсуждаются аспекты бесполого размножения у голотурий.Голотурии важны как объекты рыболовства и аквакультуры и имеют высокую коммерческую ценность. Последний обзор по бесполому размножению голотурий был опубликован 18 лет назад и включал всего 8 видов. Анализ доступной литературы показывает, что бесполое размножение в настоящее время подтверждено у 16 видов голотурий. Пять дополнительных видов также, скорее всего, способны к делению. Недавнее открытие новых видов делящихся голотурий указывает на то, что этот способ размножения более широко распространен у Holothuroidea, чем считалось ранее.Описаны новые данные об истории открытия бесполого размножения у голотурий, особенностях деления и регенерации переднего и заднего фрагментов. Бесполое размножение, очевидно, контролируется целостными системами организма, прежде всего нервной системой. По-видимому, особые молекулярные механизмы определяют место, где происходит деление вдоль передне-задней оси тела. Изменение прочности соединительной ткани стенки тела может играть важную роль при делении голотурий.Основным механизмом деления является взаимодействие матриксных металлопротеиназ, их ингибиторов и ферментов, образующих сшивающие комплексы между фибриллами коллагена. Обсуждается динамика популяций делящихся голотурий.
1. Введение
Бесполое размножение является наиболее древним способом размножения и наблюдается у представителей всех типов современных Metazoa [1–3]. Поскольку бесполое размножение тесно связано со строением животного, его типы столь же разнообразны, как и сами животные [4].Разнообразие проявлений этого явления еще больше, поскольку бесполое размножение у разных видов имеет разные биологические функции, такие как рост популяции, регуляция размеров тела, заселение новых участков, выживание в неблагоприятных условиях. Эволюция многоклеточных организмов, по-видимому, прошла через неоднократные утраты и восстановления различных форм бесполого размножения [3]. Среди современных групп беспозвоночных, размножающихся бесполым путем, особого внимания заслуживают голотурии из-за их промысловой ценности.
Голотурии, или морские огурцы, относятся к типу иглокожих. Голотурии имеют удлиненное, часто червеобразное тело, покрытое различными выростами. Как и все другие иглокожие, голотурии являются исключительно морскими животными и населяют все океаны в широком диапазоне глубин, от мелководных приливных зон до 5000 м и более. Большинство голотурий — донные организмы [5, 6], хотя есть виды, плавающие и, скорее всего, полностью пелагические [7].
Голотурии важны для коммерческого рыболовства и аквакультуры.Около 66 видов голотурий широко эксплуатируются во всем мире [8–11]. Жители этих регионов считают голотурий не только традиционным товаром, но и коммерческим ресурсом [9, 12]. Мировой отлов диких животных и аквакультурная продукция голотурий за последние 30 лет увеличились в 7 раз и составили более 20000 т сухого веса/год [12]. Кроме того, голотурии являются основным источником биологически активных веществ в биотехнологии и медицине [8, 13–16]. Они содержат широкий спектр витаминов, минералов, сапонинов, хондроитинсульфатов, полисахаридов, стеролов, фенолов, лектинов, пептидов, гликопротеинов, гликосфинголипидов и незаменимых жирных кислот [13].Таким образом, активно изучаются различные аспекты биохимии, физиологии и биологии развития этих животных.
Голотурии также примечательны тем, что обладают разнообразными способностями к регенерации [17–19]. Некоторые виды могут извергать свои внутренние органы, в основном пищеварительную систему, в ответ на различные раздражители, а затем быстро их восстанавливать [20–24]. Кроме того, многие голотурии способны регенерировать после поперечного разреза [25–28].
Некоторые виды голотурий способны к бесполому размножению.Большинство делящихся голотурий живут в тропической и субтропической зонах. Единственными исключениями являются Ocnus planci и O. lactea , деление которых наблюдалось у побережья Ла-Манша, Франция [22]. В южном полушарии Staurothyone inconspicua также встречается за пределами субтропической зоны. Этот вид с, вероятно, делящейся молодью был собран в заливе Опоссум на юге Тасмании [29]. Из-за высокой коммерческой ценности голотурий исследователи пытаются использовать их регенеративное свойство и способность к делению для разработки методов культивирования и увеличения естественных популяций [30–36].
Последний обзор по бесполому размножению голотурий был опубликован 18 лет назад [37]. В этот обзор вошли только 8 делящихся видов. С тех пор бесполое размножение наблюдается у других видов (см., например, [25, 26, 38]). Открытие новых делящихся видов свидетельствует о том, что этот тип размножения в классе Holothuroidea более распространен, чем считалось ранее. Кроме того, получены новые данные о регенерации, динамике численности и других биологических аспектах делящихся видов.Эта информация требует систематизации. Целью настоящего обзора является анализ имеющихся данных о бесполом размножении голотурий. Все названия видов, используемые в этой статье, предоставлены в соответствии с WoRMS (Всемирным реестром морских видов).
2. История открытия бесполого размножения у голотурий
Все известные к настоящему времени раздельнородящие виды голотурий относятся к двум отрядам: Aspidochirotida и Dendrochirotida. Бесполое размножение впервые было описано у дендрохиротид.Dalyell (1851, цит. Monticelli [39]) наблюдал деление Ocnus (как Cucumaria ) lactea и Ocnus (как Cucumaria ) planci . Более того, согласно Монтичелли [39], Семпер описал случай деления Havelockia (как Cucumaria ) versicolor . Однако и Далиелл, и Семпер заявили только о том, что произошло деление, но не описали процесс. Chadwik (1891, цит. Crozier [40]) дал первое краткое описание деления O.планки . Впоследствии Монтичелли [39] опубликовал всестороннее исследование бесполого размножения у этого вида.
Benham [41], по-видимому, был первым, кто привел доказательства бесполого размножения голотурий из отряда Aspidochirotida. Он наблюдал четыре экземпляра Holothuria difficilis (как Actinopyga parvula ), которые имели более светлые задние области и менее развитые трубчатые ножки в коллекции фиксированных животных с островов Кермадек. Эти животные регенерировали свои задние отделы, как полагал Бенхам [41].
Crozier [40] был первым, кто непосредственно наблюдал деление аспидохиротид. Он описал процесс деления у Holothuria surinamensis и Holothuria parvula (как Captiva ). Deichmann [42] впоследствии подтвердил бесполое размножение у H. difficilis (как Actinopyga parvula ).
Другой вид делящихся голотурий был отмечен только примерно через 40 лет. Bonham и Held [43] предоставили данные и фотографию деления Holothuria atra у Маршалловых островов.Харриотт [44] затем наблюдал в поле особей Stichopus horrens , Stichopus chloronotus и Holothuria edulis , которые подверглись делению и находились в процессе деления. Экземпляры S. horrens , описанные Harriott [44], скорее всего, принадлежали виду Stichopus monotuberculatus [45].
В статье, описывающей неотип Ocnus brunneus , Маккензи [46] указал, что этот вид дендрохиротид был способен к делению.Однако впоследствии статус этого вида не был подтвержден, и изученные особи, скорее всего, принадлежали к O. planci [47]. Впоследствии O’Loughlin и O’Hara [48] открыли первый вид делящихся дендрохиротид в 20 веке. Они сообщили, что новый вид голотурий, Squamocnus aureoruber , из прибрежных вод Австралии был способен к бесполому размножению. Конанд и др. [49] дали первое описание бесполого размножения аспидохиротиды Holothuria leucospilota , широко распространенной в тропическом Индо-Пацифике.
Дополнительные делящиеся голотурии были описаны в 21 веке. Первые четыре вида относятся к отряду Aspidochirotida. Sonnenholzner [50] сообщил о бесполом размножении у Holothuria theeli , встречающегося вдоль побережья Эквадора. Данные о способности личинок трепанга Parastichopus californicus к почкованию были также опубликованы в том же году Eaves и Palmer [51]. Это единственный вид голотурий, для которого определенно установлено бесполое размножение личинками.Могут быть и другие подобные виды [52]. Затем японские исследователи наблюдали особь Stichopus horrens в процессе деления у острова Окинава [53]. Однако, как Бирн и соавт. [45] считают, что S. horrens не способна к делению, а голотурия, наблюдаемая Kohtsuka et al. [53] было Stichopus naso . Затем способность к бесполому размножению была установлена у Holothuria hilla [38, 54]. Кроме того, образец Holothuria portovallartensis с растущим передним концом был описан Uthicke et al.[55], что, скорее всего, свидетельствует о способности этого вида к бесполому размножению.
Еще три вида расщепляющихся голотурий относятся к отряду Dendrochirotida. Это австралийский Cucuvitrum rowei [29], а также Cladolabes schmeltzii и Colochirus robustus [25, 26], которые встречаются вдоль побережья Вьетнама. Кроме того, молодь Staurothyone inconspicua может обладать способностью к поперечному делению [29].
Таким образом, в настоящее время бесполое размножение подтверждено у 16 видов голотурий (см. табл. 1). Еще пять видов, Havelockia versicolor , Staurothyone inconspicua , Stichopus monotuberculatus , S. naso и Holothuria portovallartensis , также могут быть необходимы для установления этого деления, но дальнейшие исследования могут также быть способны установить это деление. Более того, некоторые виды голотурий, скорее всего, размножаются бесполым путем на личиночной стадии [52].
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| б-почкованием; в – деление перетяжкой; е — самка; фр – фрагментация; м — самец; п — нет данных; s – деление растяжением; t – деление закручиванием. 1 Dalyell, 1851, цит. Monticelli [39]; 2 Каменев и Долматов, неопубликованные данные; 3 Semper (цит. по Monticelli [39]). | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
3. Бесполое размножение на личиночной стадии
В настоящее время имеются данные только для одного вида голотурий, P. californicus , который может размножаться бесполым путем на личиночной стадии [51]. У этого вида делению подверглось 12,2% искусственно выращенных личинок.Процесс бесполого размножения начинается на стадии долиолярии. Личинки сжимаются вокруг предпоследнего ресничного тяжа. Впоследствии углубление перетяжки на заднем конце вызывает образование почки. Эта почка (дочерний организм) сохраняет ресничную полоску и остается прикрепленной к материнской особи даже после заселения. Почка отделяется на стадии пентатактулы, и развивается нормальная аурикулярия.
Во Флоридском течении системы Гольфстрим Бальсер [52] наблюдал аурикулярии неизвестного вида голотурий, обладавшие боковыми лопастями.При искусственном культивировании боковые лопасти этих аурикулярий отделялись от особи и образовывали бластулоподобные личинки. В бластоцеле была одна «гиалиновая сфера». Личинки начинали гаструляцию, но затем дальнейшее развитие прекращалось, и животные погибали [52].
4. Бесполое размножение у взрослых
Бесполое размножение у взрослых голотурий происходит в виде поперечного деления (архитомии) и фрагментации. Наиболее подробно процесс деления описан Монтичелли [39], который выделил три механизма деления О.planci : сужением, скручиванием или растяжением. Анализ имеющихся данных о делении голотурий показывает, что различные виды голотурий используют различные комбинации методов, описанных Монтичелли (см. табл. 1). Большинство голотурий, у которых наблюдался процесс деления, начинали с образования перетяжки. Деление на осколки происходит либо путем развития и углубления перетяжки [49], либо в результате растяжения и скручивания в месте деления. Обычно задние отделы тела прикрепляются к субстрату ножками трубки, тогда как передние отделы движутся вперед или закручиваются (рис. 1) [26, 56].Закрытие раны в результате деления не описано, но, вероятно, происходит в результате сокращения кольцевых мышц стенки тела.
Продолжительность процесса варьирует от нескольких минут, как у S. chloronotus [56], до 1–5 дней, как у H. surinamensis и Cladolabes schmeltzii [26, 40]. Продолжительность деления, скорее всего, зависит от интенсивности преобразования внеклеточного матрикса стенки тела. Например, стенка кузова S.chloronotus может быстро размягчаться (J. Lawrence, личное сообщение). В отличие от других голотурий, стимуляция аутотомии хлоридом калия у этого вида вызывает быстрое размягчение и разрыв стенки тела на большой площади и выброс органов через образовавшееся большое отверстие [26].
Расположение перетяжки, по-видимому, является видоспецифическим признаком (см. табл. 1) [57]. У большинства исследованных видов голотурий деление происходит примерно посередине тела [26, 29, 40, 48, 58–60].У H. atra тело разделено на два фрагмента с соотношением длины передняя : задняя часть 4 : 5 [61]. Согласно Конанду и соавт. [49], у H. leucospilota место перетяжки смещено к переднему концу, а длина передних фрагментов составляет примерно 19% от общей длины тела. У Colochirus robustus соотношение разделенных частей тела составляет 2 : 1; передний фрагмент вдвое длиннее заднего [25].
Некоторые виды голотурий могут разделяться на несколько частей тела одновременно. Это давно известно для видов O. lactea и O. planci , как описано Dalyell (1851, цит. Monticelli [39]) и Monticelli [39]. Cucuvitrum rowei также подвергается фрагментации [29]. Повторное деление на фрагменты, не регенерирующие полностью утраченный участок, процесс, сходный с фрагментацией, происходит у аспидохиротиды H.parvula [60] и дендрохиротид Cladolabes schmeltzii [25].
5. Механизмы деления
Бесполое размножение у голотурий — очень сложный процесс, в котором задействованы различные механизмы и системы органов. Во-первых, деление сопровождается сложным поведением, таким как растяжение и скручивание в месте деления. Более того, согласно Пурвати [57], голотурии перестают питаться до деления. Такое сложное поведение, очевидно, контролируется целостными системами организма, в первую очередь нервной системой.Это согласуется с тем, что аутотомия, процесс, близкий к делению, нервно опосредован [65, 66]. Во-вторых, деление происходит на определенных участках тела, что является видоспецифическим признаком. Это означает, что особые молекулярные механизмы определяют расположение деления вдоль передне-задней оси тела. В-третьих, стенка тела, или дерма, у голотурий состоит почти исключительно из соединительной ткани [19, 67]. Поэтому деление тела делением невозможно без преобразования внеклеточного матрикса.
В настоящее время исследований вклада нервной системы в регуляцию деления и изучения клеточных и молекулярных механизмов деления у голотурий не проводилось. Имеются только данные о свойствах внеклеточного матрикса. Соединительная ткань голотурий обладает уникальной способностью изменять свои механические свойства под действием различных факторов [68, 69]. Поэтому ее называют мутабельной коллагеновой тканью (МСТ) [70] или уловкой соединительной ткани [71]. Это свойство, по-видимому, очень важно для деления [72–75].
Соединительная ткань дермы у голотурий может находиться в трех состояниях: жестком, стандартном и мягком [74]. Механическое растяжение участка дермы приводит к уменьшению жесткости соединительной ткани и способствует ее переходу в мягкое состояние. Предложен механизм деформационного размягчения при аутотомии у голотурий [74]. Вначале размягчается небольшой участок дермы в области будущего разрыва. Затем мышцы сокращаются, и давление в целомической полости повышается.Повышенное давление вызывает обширную деформацию в размягченной части. Жесткость резко снижается по мере превышения порогового значения растяжения. Одновременно регистрируется увеличение величины диссипации (выделения энергии системой), что, скорее всего, свидетельствует о разрыве межмолекулярных связей. Это рассеивание позволяет дерме продолжать деформироваться при одинаковом давлении (или при более низких значениях). Таким образом, возникает положительная обратная связь: чем больше деформируется дерма, тем легче она деформируется.
На клеточном уровне эти свойства МСТ определяются особым типом клеток – околосвязочными клетками соединительной ткани [68]. Цитоплазма юксталигаментарных клеток содержит многочисленные гранулы, окруженные мембраной. Считается, что эти клетки секретируют вещества, полученные из гранул, в ответ на сигналы нервной системы. Эти вещества влияют на взаимодействие между молекулами внеклеточного матрикса соединительной ткани, вызывая тем самым быстрые изменения механических свойств последней [76–81].
Иммуноцитохимические методы выявили два белка в гранулах юксталигаментарных клеток, тензилин и стипарин, которые, по-видимому, участвуют в механизмах изменения свойств МСТ (Keene, Trotter, неопубликованные данные, цит. по Wilkie [69]). Эти белки связываются с фибриллами коллагена и, как полагают, образуют поперечные связи между ними in vivo , таким образом стабилизируя волокна коллагена (т.е. пучки фибрилл). Нуклеотидная последовательность транскрипта гена тензилина и предсказанная аминокислотная последовательность тензилина в настоящее время известны для одного вида голотурий Cucumaria frondosa [82].Этот белок проявляет высокую гомологию с тканевыми ингибиторами металлопротеиназ (ТИМП), что свидетельствует об участии матриксных металлопротеиназ (ММП) в функционировании МСТ [69]. Исследования связок компаса-депрессора морского ежа Paracentrotus lividus выявили некоторые протеазы, обладающие желатиназной активностью [83]. Блокирование их специфическим ингибитором увеличивает жесткость связок. Согласно предложенной модели, жесткость МСТ зависит от взаимодействия между тремя белковыми группами, ММП, ТИМП и комплексами поперечных связей, соединяющими коллагеновые фибриллы друг с другом [83].При увеличении высвобождения ТИМП блокируется ММП, развиваются поперечные связи между фибриллами коллагена и усиливается МСТ. Возрастающая концентрация или активность ММП во внеклеточном матриксе вызывает разрушение комплексов поперечных связей. Это разрушение позволяет фибриллам коллагена скользить друг относительно друга, что приводит МСТ в податливое состояние. Помимо включения в комплексы с поперечными связями, тензилин может быть эндогенным ингибитором ММР.
Некоторые внутренние органы голотурий, такие как кишечник и продольные мышечные тяжи (вместе со стенкой тела), разрываются во время деления.Аутотомия этих органов у Eupentacta quinquesemita происходит в результате полной потери растяжимости их соединительной ткани [84]. Этому процессу способствует сокращение мышц. Изменения в органах, по-видимому, начинаются с разрыва целомического эпителия. Затем соединительная ткань инфильтрируется целомической жидкостью. Предполагается, что целомическая жидкость голотурий содержит фактор эвисцерации, который воздействует на соединительную ткань, провоцируя потерю ее растяжимости [66]. В отличие от других иглокожих аутотомия (потрошение) у голотурий может происходить без участия юксталигаментарных клеток [84].Отростки этих клеток оставались в значительной степени интактными, несмотря на обширное разрушение внутренних органов.
Аналогичный механизм, который изменяет жесткость соединительной ткани дермы, может действовать и при бесполом размножении голотурий. При делении происходит локальное размягчение дермы из-за активности ММР в определенном месте вдоль передне-задней оси голотурий. Этот процесс может сопровождаться сокращениями лучевых мышц стенки тела, вызывающими дополнительное растяжение.Продолжительное местное воздействие на дерму вызовет еще большее размягчение, и в стенке тела образуется перетяжка. Дополнительные скручивающие или растягивающие движения тела ускорили бы деление. Разница в продолжительности деления у разных делящихся видов, скорее всего, определяется различиями в свойствах внеклеточного матрикса и активности ММП, ТИМП и других ферментов.
6. Регенерация внутренних органов после деления
Набор органов, оставшихся в фрагментах после деления, примерно идентичен для всех исследованных видов делящихся голотурий (рис. 2(а) и 2(б)).Различия связаны только с расположением места деления. У H. atra , H. parvula , S. chloronotus , Colochirus robustus и Cladolabes schmeltzii , осуществляющих деление в средней или задней частях тела, сложные апередние фрагменты сохраняют (AC), гонады и один или два сегмента кишечника [25, 26, 58, 60, 85]. Задний фрагмент этих животных содержит клоаку, большую часть кишечника и дыхательные деревья.Задний фрагмент аспидохиротид сем. Holothuriidae также содержит кювьеровы трубочки. У H. leucospilota , который делится ближе к переднему концу, гонады могут оставаться в заднем [49] или переднем фрагменте [57].
Возможны два варианта деления кишечника, которые, скорее всего, зависят от положения кишечника в полости тела в момент деления [86]. Если кишечные петли расположены кзади от места деления, разрыв происходит только в середине первого нисходящего отдела кишки (1ДП).Если кишечные петли пересекают плоскость деления, пищеварительная трубка делится на три места, и в обоих отделах животного остается по два фрагмента.
6.1. Макроморфологические особенности регенерации
Регенерация внутренних органов после деления с разной степенью подробности описана для 6 видов голотурий: H. difficilis [42], H. parvula [60], H. atra [42]. 85, 86], H. leucospilota [49], S. chloronotus [58] и C.schmeltzii [26, 87]. Все описания основаны только на анализе фрагментов, собранных в природе, и, таким образом, продолжительность регенерации остается неизвестной. Все данные предполагают, что регенерация внутренних органов после деления одинакова у всех голотурий, по крайней мере, на макроморфологическом уровне (рис. 2).
В переднем фрагменте регенерация после деления начинается с частичной атрофии поврежденной 1ДП, длина которой соответственно уменьшается. Одновременно ушивают рану на заднем конце.Более того, изолированный фрагмент кишки, если он есть, расщепляется. Затем конец 1DP начинает расти назад, вниз по брыжейке, образуя зачаток кишечника. Одновременно на заднем конце переднего фрагмента развивается клоака. Зачаток кишечника удлиняется и врастает в клоаку, восстанавливается целостность пищеварительной системы (рис. 2, в). Животные на этой стадии регенерации восстанавливают концевые отделы продольных мышечных тяжей (LMB), которые были повреждены при делении.Концы ЛМБ истончаются и растут в сторону клоаки. После слияния кишечника и клоаки на дорсальной стороне клоаки появляются зачатки дыхательных деревьев (рис. 2(г)).
На этом этапе также начинается рост тела. На заднем конце появляется небольшой вырост, который впоследствии удлиняется, образуя задний отдел животного (рис. 2(д)). Дыхательные деревья развиваются с ростом заднего конца. Дыхательные деревья постепенно разрастаются и образуют боковые ветви.У этих животных терминальные области LMB остаются тоньше, чем обычно. Восстановление БКМ, скорее всего, будет завершено позже.
Основным событием в заднем фрагменте является регенерация АК. Сначала на переднем конце животного между разорванными концами амбулакров образуется соединительнотканное вздутие, представляющее собой зачаток АК (рис. 2(ж)). Затем в зачаток АК врастают терминальные отделы лучевых нервных тяжей и радиальных водно-сосудистых каналов. В последующем эти участки образуют соответственно нервное кольцо и циркулярный водно-сосудистый канал вокруг АК.Разорванный передний отдел кишки трансформируется по мере развития АК. В месте повреждения кишка истончается и начинает расти вверх по брыжейке (рис. 2(ж)). Зачаток кишечника в виде тонкой трубки врастает в АК, и целостность пищеварительной системы восстанавливается (рис. 2(з)). Затем животное начинает расти. На переднем конце появляется небольшой вырост, который постепенно удлиняется, образуя передний отдел животного (рис. 2(и)).
6.2. Микроанатомические особенности регенерации
Регенерация после деления на клеточном уровне описана только для C. schmeltzii [87]. Здесь отличительной особенностью развития пищеварительной системы обоих фрагментов является образование кишечника из двух зачатков. Энтодермальная область (кишка) образуется в результате дедифференцировки и миграции энтероцитов оставшихся 1ДП. При дедифференцировке энтероциты теряют много секреторных гранул и микроворсинок, а их высота уменьшается.Тем не менее, энтероциты сохраняют свои межклеточные соединения. Просветный эпителий кишечника не разрушен. После слияния с клоакой (передний фрагмент) или АК (задний фрагмент) и восстановления целостности пищеварительного тракта энтероциты становятся специализированными в зависимости от их положения вдоль кишечника [87]. В целом механизмы регенерации кишечника у C. schmeltzii при бесполом размножении аналогичны механизмам регенерации кишечника после аутотомии у других видов голотурий [18, 21, 23, 88–91].В обоих случаях утраченные участки образуются за счет дедифференцировки и миграции оставшихся клеток люминального эпителия.
Эктодермальные отделы (глотка, пищевод и задний конец клоаки), по-видимому, образуются из клеток эпидермиса, которые мигрируют из эпидермиса стенки тела животного в соединительную ткань стенки тела и зачатка АК. В этом случае клетки, скорее всего, сохраняют связи друг с другом, а их межклеточные контакты не нарушаются.На ранних стадиях регенерации кишечника эти клетки начинают синтезировать кутикулу. В дальнейшем эктодермальная область сливается с энтодермальной областью; таким образом, целостность пищеварительной системы восстанавливается.
Дыхательные деревья в переднем фрагменте развиваются за счет трансформации дорсальной стенки переднего отдела клоаки. Отличительной особенностью регенерации дыхательной системы является быстрая специализация клеток просветного эпителия. Эти клетки имеют ламеллы на апикальной поверхности и связаны между собой комплексом специфических межклеточных контактов на ранних стадиях формирования.Поэтому регенерация дыхательных деревьев у C. schmeltzii сходна с развитием и регенерацией этих органов после эвисцерации у Apostichopus japonicus [92, 93].
Новые мышечные пучки формируются в регенерированных концах ЛМБ из целомического эпителия, покрывающего мышцы. Сначала в соединительную ткань встраиваются группы клеток. После этого эти клетки трансформируются в миоциты, а в их цитоплазме наблюдаются миофиламенты.Эти группы клеток затем отделяются от эпителия и образуют новые мышечные пучки. Этот процесс идентичен росту и регенерации мышц у иглокожих [94–98].
Регенерация АК в заднем фрагменте также происходит из клеток остальных органов [87] и аналогична репарации после эвисцерации [18, 99]. Клетки терминальных сегментов лучевых водно-сосудистых каналов и радиальных нервных тяжей дедифференцированы и мигрируют вниз по соединительнотканной зачатке АК.В миоэпителиальных клетках просветного эпителия водно-сосудистых каналов нарушен сократительный аппарат. Миофиламенты собираются в веретенообразные структуры и выбрасываются из цитоплазмы. Межклеточные соединения при дедифференцировке не нарушаются, и клетки мигрируют внутри эпителия. Поэтому трубчатый зачаток водно-сосудистого канала формируется и постепенно растет по ходу АК. Нервные тяжи, по-видимому, растут аналогичным образом. И нейроны, и глиальные клетки участвуют в регенерации нервной системы.
В целом регенерация внутренних органов после деления у C. schmeltzii аналогична регенерации этих структур после потрошения или искусственного повреждения. Большую роль в восстановлении играет трансформация остальных отделов органов. Основными механизмами являются дедифференцировка и перемещение слоев эпителия (эпителиальный морфогенез).
6.3. Рост тела
Рост тела начинается с формирования внутренних органов (рис. 2(e) и 2(i)).На этой стадии у большинства голотурий появляются начальные признаки регенерации. В месте деления дерма депигментируется и образуется бугорок. Продолжительность роста тела варьирует в широких пределах и, по-видимому, зависит от вида. У особей C. schmeltzii , которые, скорее всего, были отловлены вскоре после деления и не имели визуальных признаков роста на конце, в течение 25 дней при животных содержали в искусственных условиях (Долматов, неопубликованные данные).У S. chloronotus для роста внешней области до нормальных размеров требовалось до одного месяца [59, 64]. Согласно Jaquemet et al. [100] Регенерация H. atra после деления заняла около шести месяцев.
Оценки скорости деления и регенерации обычно основаны на внешней морфологии животных, особенно на наличии растущего переднего или заднего конца тела. При этом все животные, не проявившие отчетливого нароста в месте деления, объединялись в одну группу и рассматривались как просто разделившиеся особи.Эти голотурии использовались для оценки интенсивности деления [49, 57, 85, 86]. Но на самом деле в эту группу входят голотурии на разных стадиях регенерации [60, 85, 87]. Продолжительность этих стадий в природе неизвестна, и оценки у голотурий могут быть только косвенными и основанными на экспериментах по регенерации после искусственной рубки. Можно предположить, что развитие органов при бесполом размножении и после искусственного вырезания идет с одинаковой скоростью. У C. schmeltzii формирование внутренних органов без роста наружной области происходит примерно за 30 дней [26].Следовательно, у животных, имеющих небольшой вырост на заднем конце, деление произошло более месяца назад. Продолжительность регенерации внутренних органов у других голотурий может составлять от 1,5 до 3 месяцев [26, 33, 35]. Таким образом, животные с небольшими выростами могли разделиться еще несколько месяцев назад. Этот факт следует учитывать при оценке сезонной интенсивности деления.
6.4. Развитие гонад
Развитие гонад является важным вопросом в изучении бесполого размножения голотурий, так как один из фрагментов после деления полностью утрачивает репродуктивную систему, включая набор первичных половых клеток.Регенерация гонад, по-видимому, происходит у всех раздельнородящих голотурий, так как даже популяции с высокой степенью деления продолжают размножаться половым путем, а доля бесполых особей не увеличивается [59, 60, 64].
Развитие половой системы после деления изучено только у одного вида H. parvula [27]. Половые железы формируются поздно. Зачатки половой системы наблюдались только у тех особей, у которых полностью сформировались остальные органы.Основание гонады развивается сначала в виде одного или нескольких скоплений клеток в брыжейке кишечника. Гистологический анализ показал, что в основании гонады содержатся первичные половые клетки. Затем гонадные канальцы начинают расти от основания гонады. В процессе развития гонодук начинает формироваться и растет от гонады к дорсальной области стенки тела.
7. Искусственное деление
Особого внимания заслуживает так называемое «искусственное деление». «Искусственное деление» — разделение голотурии на две части путем стягивания тела резиновой лентой [30, 32, 34–36] или путем поперечного разрезания [31].Монтичелли [39] провел первые такие эксперименты. Искусственное деление голотурии на две части весьма далеко от естественной стимуляции бесполого размножения и может рассматриваться лишь как имитация деления. При использовании резинки поведенческие реакции, связанные с делением (сжатие, скручивание, растяжение), отсутствовали [32]. Скорее всего, никакие внутренние механизмы, запускающие деление, не были задействованы. Простое разделывание животного имеет еще меньше общего с естественным делением.Тем не менее, такие исследования важны и полезны, чтобы узнать больше о способностях к регенерации для разработки технологий культивирования и увеличения популяции голотурий. Эксперименты с поперечным разрезанием показали, что не все голотурии могут восстанавливать оба фрагмента тела [18, 26, 28, 35, 36]. Например, передний и задний фрагменты Actinopyga mauritiana погибают после перетяжки тела резинкой [35]. У Holothuria fuscogilva , A. miliaris и Stichopus variegatus в цельных животных могут регенерировать только задние отделы [35].С другой стороны, более мелкие (молодые) особи A. mauritiana , H. fuscogilva и S. variegatus имеют более высокую выживаемость и более короткое время регенерации по сравнению со взрослыми особями [36]. После поперечного разрезания оба фрагмента Holothuria pervicax , H. impatiens и Massinium magnum погибают [25, 26]. Только задние отделы Apostichopus japonicus , Holothuria scabra , Ohshimella ehrenbergi и Colochirus quadrangularis способны регенерировать утраченные передние структуры после такой операции [18, 25, 26].
8. Динамика населения
8.1. Сезонные особенности и связь с половым размножением
Бесполое размножение голотурий может происходить в течение всего года, при этом активность варьируется в зависимости от популяции и сезона. В популяциях H. atra [44, 64, 101–103], S. chloronotus [59], H. edulis [64] и H. difficilis [63], обитающих в южных полушарии наибольшая активность деления наблюдалась зимой, когда температура воды была самой низкой.Пик полового размножения у этих видов приходится на теплый период года (ноябрь-февраль). В северном полушарии наибольшая активность деления регистрировалась в летние месяцы и совпадала с периодом полового размножения [60, 62, 104]. Эти различия в сезоне деления между северным и южным полушариями могут быть вызваны различиями во времени полуденных отливов летом и зимой [62].
Очевидно, что бесполое размножение отрицательно влияет на половую активность.Половина животных после деления (главным образом задний фрагмент) не имеют гонад. Соответственно уменьшилась доля половых особей в популяциях даже тех видов, у которых оба типа размножения были очень синхронизированы и в разное время года [59, 60]. Типичным примером негативного влияния деления на половое размножение является популяция H. atra вдоль побережья Тайваня. Поскольку сезон полового размножения совпадает с сезоном максимального деления, две половинки, образующиеся в результате деления, не имеют гонад и не нерестятся [62, 104].У H. parvula доля особей со зрелыми гонадами в течение года не превышает 10% [60].
Снижение репродуктивного потенциала проявляется не только низким гонадным индексом, но и смещенным соотношением полов с преобладанием самцов и меньшими размерами тела. Для некоторых популяций раздельнородящих голотурий характерно смещение в соотношении полов [39, 44, 59, 63, 105, 106]. Таким образом, соотношение мужчин и женщин может достигать 31 : 1 [59]. Считается, что бесполое размножение провоцирует смещенное соотношение полов у иглокожих [106–108].Согласно Макговерну [109], одной из основных причин этого явления является различие в скорости деления самцов и самок. Однако эта связь не так однозначна, как может показаться. Существуют также популяции делящихся видов, в которых соотношение полов составляет 1 : 1 [59]. Более того, соотношение полов может меняться с возрастом. По данным Harriott [44], среди H. atra особей, весивших менее 100 г, преобладали самцы (8,5 : 1). Доля самок увеличивалась по мере увеличения массы тела.Соотношение самцов и самок среди животных массой более 1000 г составляло 0,7 : 1. В популяции H. parvula на Бермудских островах систематическая ошибка соотношения полов, по-видимому, отсутствовала [60]. В популяции Cladolabes schmeltzii из залива Нячанг самок было больше, чем самцов (Каменев, Долматов, неопубликованные данные). Существует множество причин предвзятого соотношения полов, которые могут различаться между видами и популяциями [109–112]. Однако специальных исследований причин смещения соотношения полов у голотурий не проводилось.Утик и др. [106] отметили, что уменьшение доли самок может быть результатом более высокой смертности среди взрослых самок, более высокой скорости деления среди самцов, более низкого полового пополнения или более высокой смертности среди личиночных самок.
Высокая интенсивность деления вызывает постепенное снижение средней массы тела и размеров [39, 59, 75]. Дальнейшая судьба популяции, по-видимому, зависит от скорости роста фрагментов и условий среды [102]. Согласно Монтичелли [39], повторное деление в O.planci приводил к уменьшению размеров тела человека и смерти. Средняя масса особей S. chloronotus в некоторых популяциях на Реюньоне за три года наблюдений снизилась в 1,5 раза (с 55 до 37 г) [59]. Однако, как было показано в популяциях Большого Барьерного рифа, масса тела может восстановиться до нормы в течение нескольких месяцев [75].
8.2. Популяционная генетика
Очень мало известно о генетической структуре популяций расщепляющихся голотурий.Строение изучено только у двух видов: H. atra [105, 113] и S. chloronotus [106, 113, 114]. Бесполое размножение оказывает значительное влияние на генетическую структуру популяций этих видов [114]. Генетическое разнообразие у делящихся голотурий снижено. В наиболее изученной популяции 40–60% особей произошли в результате бесполого размножения [105, 106, 113]. В некоторых популяциях S. chloronotus , почти полностью состоящих из самцов, только до 20% всех особей размножались половым путем [114].«Генетическая связь» между популяциями происходила только за счет личинок, размножающихся половым путем. Клоны голотурий ограничены местными популяциями [114].
9. Факторы, влияющие на бесполое размножение
Интенсивность бесполого размножения сильно различается между популяциями одного и того же вида голотурий [59, 75]. Это указывает на то, что окружающая среда играет важную роль в запуске и регулировании деления [59]. Факторы, влияющие на бесполое размножение личинок голотурий, неизвестны.Тем не менее имеются экспериментальные данные на других иглокожих, которые служат основанием для двух предполагаемых механизмов стимуляции бесполого размножения. Личинки морских звезд и морских ежей при культивировании при оптимальной температуре и разнообразной кормовой базе клонируются с большей скоростью [115, 116]. Скорее всего, личинки, находившиеся в наиболее подходящих условиях обитания, были стимулированы к бесполому размножению. Рост численности особей, живущих в оптимальных условиях, по-видимому, увеличивает вероятность успешного развития, метаморфоза и достижения ювенильной стадии, что в конечном итоге приводит к росту численности популяции.
Однако у морского ежа Dendraster excentricus скорость бесполого размножения у личинок pluteus увеличивается при введении в среду культивирования внешней слизи рыб (сигналы хищника) [117–119]. Такое поведение является реакцией избегания хищников. Воздействие слизи активизирует почкование и деление личинок. Поэтому размеры тела уменьшаются, что позволяет особям более успешно убегать от хищников [118].
У взрослых голотурий основными факторами, влияющими на скорость бесполого размножения, являются низкая экологическая устойчивость, высокая смертность, малые размеры тела и низкая половая репродуктивная активность [37, 75, 102, 103].Эмсон и Уилки [22] отметили, что многие делящиеся виды хрупких звезд обитают в приливных или мелководных водах. В этой среде офиуры подвергаются воздействию сильно различающихся факторов окружающей среды, которые могут вызвать бесполое размножение. У Colochirus robustus деление, скорее всего, стимулируется стрессом при содержании в неподходящих условиях [25].
Влияние окружающей среды на интенсивность деления также было продемонстрировано для H. atra . Было замечено, что у этого вида есть две размерные морфы.Мелкие особи могут размножаться как половым, так и бесполым путем, тогда как крупные размножаются только половым путем [62, 63, 104]. Оба размера наблюдаются у одного и того же вида и представляют собой фенотипические экотипы [11]. Мелкие расщепляющиеся особи обитают в литорали, для которой характерны значительные колебания условий среды, тогда как крупные приспособлены к более стабильной сублиторальной зоне [62, 75, 85, 102, 104, 105, 120]. При перемещении мелких особей из литорали в сублиторали они становятся крупными [102, 104].Более того, более высокая доступность пищи из-за уменьшения численности популяции стимулирует рост особей и останавливает бесполое размножение [102, 103].
Размер некоторых популяций делящихся голотурий, несмотря на высокую скорость деления, может длительное время оставаться постоянным [44, 75, 85, 101–104, 120, 121]. Этот сценарий указывает на то, что часть особей элиминируется из этой популяции из-за гибели при бесполом размножении или эмиграции в другие местообитания [44, 85, 102]. Согласно модели, предложенной Утиком [75], стабильность численности популяции указывает на наличие механизма обратной связи между бесполым размножением и смертностью и эмиграцией.Этот механизм поддерживает плотность популяции вблизи средних значений, которые реализуют потенциал среды.
10. Заключение
Анализ доступной литературы показывает, что бесполое размножение описано для 16 видов голотурий (см. табл. 1). Пять дополнительных видов также, скорее всего, способны к делению, но для подтверждения этого необходимы дополнительные исследования. Более того, некоторые виды голотурий могут размножаться бесполым путем на личиночной стадии [52]. Недавнее открытие новых видов делящихся голотурий указывает на то, что этот способ размножения более широко распространен у Holothuroidea, чем считалось ранее.
Бесспорно, бесполое размножение играет большую роль в жизнедеятельности голотурий и поддерживает численность популяции. Особое значение деление приобретает для промысловых видов, подвергающихся массовому перелову [12]. Активное деление позволяет голотуриям поддерживать большую популяцию и смягчать негативные внешние воздействия. Однако, несмотря на некоторые преимущества при определенных условиях, клонирование, скорее всего, выгодно только в краткосрочной перспективе. Уменьшение размера тела и активности полового размножения в течение длительного времени может уменьшить размер популяции и выживаемость вида.В отличие от других клональных животных, таких как книдарии, развитие внутренних органов и дальнейший рост у голотурий могут быть длительными (в течение нескольких месяцев). Гонада формируется только через несколько месяцев после деления. Поэтому снижается интенсивность полового размножения и, скорее всего, возникает перекос в соотношении полов.
Очевидно, что адаптивное значение и преимущества бесполого размножения у голотурий могут быть связаны с его сочетанием с половым размножением. Наличие двух экоморфов является заметной адаптацией, поскольку позволяет животным использовать совершенно разные условия обитания.Более того, бесполое размножение происходит как механизм поддержания больших размеров популяций, где эмиграция личинок значительна [75, 122].
Многие проблемы, касающиеся бесполого размножения голотурий, еще предстоит решить. В частности, отсутствуют исследования клеточных и молекулярных механизмов деления. В настоящее время остается неизвестным, какие факторы (гены) определяют местонахождение участка деления тела голотурии. Вопросы восстановления репродуктивной системы остаются неизученными.Источник первичных половых клеток во фрагментах, лишенных гонад, также неясен. Кроме того, необходимы дополнительные исследования факторов, стимулирующих деление и повышающих активность бесполого размножения в популяции.
Бесполое размножение у голотурий — примечательное и малоизученное явление. Изучение деления и связанной с ним регенерации необходимо для понимания размножения голотурий. Не исключено, что разработка методов стимуляции бесполого размножения или регенерации после искусственной обрезки поможет восстановить популяцию голотурий и даст дополнительный экономический эффект.
Конфликт интересов
Автор заявляет об отсутствии конфликта интересов в связи с публикацией данной статьи.
Благодарности
Автор благодарен Джону Лоуренсу за критическое прочтение статьи. Большое спасибо рецензентам за полезные комментарии. Работа выполнена при частичной поддержке РФФИ (грант № 14-04-00239) и Правительства РФ (грант № 11.Г34.31.0010).
Несмотря на разногласия, виды рыб, не способные к половому размножению, процветают – Медицинская школа Вашингтонского университета в Сент-Луисе.Луи
Посетите центр новостейУспех бесполого размножения у амазонской молли — эволюционная аномалия
Манфред ШартльМеждународная группа ученых секвенировала геном амазонской молли, рыбы, размножающейся бесполым путем. Исследователи ожидали, что бесполый организм окажется в невыгодном генетическом положении, но амазонская молли процветает.
Считается, что очень редкие животные, размножающиеся бесполым путем — лишь примерно одно из 1000 всех ныне живущих видов позвоночных — находятся в эволюционно невыгодном положении по сравнению со своими собратьями, размножающимися половым путем.Но эта теория не верна в отношении амазонской моллинезии, полностью женского вида рыб, который тысячелетиями процветал в пресных водах вдоль мексиканско-техасской границы.
Чтобы лучше понять, как размножение этой рыбы отклоняется от нормы, международная группа ученых секвенировала геном первой амазонской моллинезии и геномы исходных родительских видов, которые создали эту уникальную рыбу. Их результаты показывают, что процветающее существование моллинезии не является полностью неожиданным, поскольку они обнаружили, что рыба имеет выносливый генетический состав, который часто редко встречается в природе и дает животным некоторые предсказанные преимущества выживания.
«Похоже, что звезды сошлись для этого вида», — сказал первый автор Уэсли С. Уоррен, доктор философии, помощник директора Института генома Макдоннелла в Медицинской школе Вашингтонского университета в Сент-Луисе. «Гибридизация геномов двух разных видов в один новый потребовала бы почти идеальной совместимости между элементами этих родительских геномов, чтобы обойти половое размножение, практикуемое большинством видов позвоночных».
Результаты исследования опубликованы 12 февраля в журнале Nature Ecology & Evolution.
С тех пор, как ученые в 1932 году определили, что амазонская молли была первым известным бесполым позвоночным, они задавались вопросом, как это произошло.
Одна из теорий, разъясняющая, почему бесполое размножение должно стоять на пути к устойчивости вида, заключается в том, что если во время размножения не будет введена новая ДНК, то вредные генные мутации могут накапливаться в последующих поколениях, что в конечном итоге приведет к вымиранию. Другая гипотеза гласит, что, поскольку бесполое размножение ограничивает генетическое разнообразие внутри вида, животные в конечном итоге становятся неспособными адаптироваться к изменениям в окружающей среде.
«Ожидается, что эти бесполые организмы находятся в невыгодном генетическом положении», — сказал Уоррен, также доцент кафедры генетики. «В природе амазонская молли чувствует себя неплохо».
Исследователи обнаружили, что амазонская моллинезия возникла в результате полового размножения двух разных видов рыб, когда атлантическая моллинезия впервые спарилась с моллинезией-парусником от 100 000 до 200 000 лет назад. С тех пор получившийся в результате амазонский молли был гибридным видом, который удивительно остался замороженным во времени эволюции, но все еще продолжает процветать.
— Это около 500 000 поколений, если считать до сегодняшнего дня, — сказал Уоррен. «Ожидается, что за это время накопится много вредных мутаций, но это не то, что мы обнаружили».
Молли амазонская размножается путем «спаривания» с самцом рыбы родственного вида. Но ДНК самца не включается в потомство. Вместо этого спаривание с рыбой-самцом запускает репликацию всего материнского генома. По сути, моллинезии клонируют себя. Они не откладывают яйца, а вместо этого рожают большие выводки живых потомков.
Ученые давно предполагают, что клоны, не избавившись от вредных мутаций, должны подвергаться распаду генома и, в конечном итоге, вымирать через поколения.
«Это исследование завершает интенсивное совместное исследование, знаменующее первое представление о геномных особенностях бесполых позвоночных и создающее платформу для будущей молекулярной, клеточной и исследовательской работы с этим интересным видом», — сказал Майкл Линч, доктор философии, директор Центр биодизайна механизмов эволюции Университета штата Аризона.
Итак, хотя амазонская молли процветала тысячи лет, она по-прежнему сопротивляется раскрытию своих геномных секретов — пока.
«Возможно, у амазонской молли есть лучшее из обоих миров», — сказал Манфред Шартль, профессор и заведующий кафедрой биохимии Вюрцбургского университета в Вюрцбурге, Германия. «Кажется, у него есть некоторые преимущества, которые мы наблюдаем у видов, размножающихся половым путем, и другие преимущества, обычно наблюдаемые у видов, производящих потомство неполовым путем, например большая численность популяции»
.Какими бы ни были причины, исследователи заявили, что амазонская молли является исключением из представлений об эволюционных недостатках бесполого размножения.
Уоррен В.С. и др. Клональный полиморфизм и высокая гетерозиготность в целибатном геноме амазонской молли. Природа, экология и эволюция, 12 февраля 2018 г.
Эта работа была поддержана Национальным институтом здравоохранения (NIH), номера грантов 2R24-OD011198-04A1, R24-OD011120 и R24-RR032658-01. Дополнительное финансирование от Национального научного фонда, номер гранта NSF DBI-1564611, Немецкого исследовательского фонда, Немецкого научного фонда, Европейской организации молекулярной биологии (EMBO), Fundacio Zoo Barcelona, Secretaria d’Universitats i Recerca del Departament d’Economia i Coneixement de la Generalitat de Catalunya, Программа внутренних исследований Национального института здоровья, Национальная медицинская библиотека, Wellcome Trust и Европейская лаборатория молекулярной биологии.
Медицинская школа Вашингтонского университета, состоящая из 2100 штатных и волонтерских врачей, также является медицинским персоналом детских больниц Барнс-Джуиш и Сент-Луис. Медицинский факультет является одним из ведущих медицинских исследовательских, учебных и лечебных учреждений в стране и в настоящее время занимает седьмое место в стране по версии US News & World Report. Медицинская школа связана с BJC HealthCare благодаря своим связям с детскими больницами Barnes-Jewish и St. Louis Children.
Половое размножение: как природа создает сложность и разнообразие
Снова пора: еще один день празднования Земного календарного года, календаря, который сжимает возраст Земли (4,5 миллиарда лет) в один календарный год.
17 сентября история эволюции достигает следующей вехи, когда «появляется» или, лучше сказать, начинается половое размножение. Около 1,3 миллиарда лет назад половое размножение начало смешивать гены и проложило путь к большому разнообразию, которое мы наблюдаем сегодня.
Это чрезвычайно важно на этой временной шкале, потому что позволяет организмам начать расчесывать гены, позволяя следующему поколению делать больше, чем его родители; повышение шансов на выживание.
На этой неделе мы отмечаем «день рождения» полового размножения в рамках нашего ежегодного празднования «Счастливого дня рождения жизни». В марте мы рассказывали вам о дне появления фотосинтеза. И, мы рассказали вам о том, как Жизнь впервые появилась на Земле в виде одноклеточного организма 25 февраля.Согласно земному календарю, где 144 года равны одной секунде, если Земля «родилась» 1 января, то 25 февраля было бы днем зарождения жизни.
Что такого важного в половом размножении?
Первые живые организмы просто воспроизводили себя как копии. Это было легко и быстро. До тех пор, пока 1,3 миллиарда лет назад не произошло что-то новое.
Секс!
Некоторым организмам сначала нужно было найти себе пару, чтобы размножаться. Это было медленно, потому что не все потомки были самками, что замедляло процесс.Итак, почему изменение? Почему эта новая идея выжила и стала такой успешной? Каковы были преимущества по сравнению с системой быстрого копирования, которая могла быстрее заполняться?
Половое размножение означает, что организм может комбинировать половину своих генов с половиной чужих генов, что создает новые комбинации для каждого поколения. Хотя половое размножение происходит медленнее, в результате потомство может делать больше, чем его родители.
Перетасовка и рекомбинация генов создают сложность, но также и больше способов выжить в меняющемся мире.Природа не статична. Если вы сможете смешать гены, у вашего потомства больше шансов выжить. Половое размножение стало успешным, потому что организмы начали изобретать больше способов находить пищу, справляться с патогенами и привлекать лучшего партнера. Генетическое разнообразие означает, что вид может меняться в течение последующих поколений.
Если дизайн никогда не меняется, возможно, он не сможет адаптироваться к новым условиям. Сложность и даже мутации, которые сейчас не работают, подобны лишним идеям в заднем кармане.Они могут не соответствовать старой системе, но когда система изменится — вуаля!
Сегодня все еще существуют организмы, которые могут воспроизводиться сами по себе (бесполым путем), такие как некоторые бактерии, растения, черви и даже некоторые ящерицы и змеи. Это сопротивление может стать тупиком, если мы столкнемся с необходимостью выживания в условиях быстрых изменений, которые сейчас переживает планета.
