
Генетическая гистосовместимость партнеров
Система HLA обеспечивает регуляцию иммунного ответа, контролируя такие важнейшие физиологические процессы, как взаимодействие иммунокомплектных клеток организма, распознавание клеток, запуск и реализация иммунного ответа. Система HLA представляет собой индивидуальный набор различного типа белковых молекул, расположенных на клеточной поверхности. Набор антигенов (HLA-статус) уникален для каждого человека.
К первому классу главного комплекса гистосовместимости относятся молекулы типов HLA-А, -В и -С. Антигены первого класса находятся на поверхности любых клеток. Представителями второго класса являются HLA-DQ, -DP, -DR. Антигены второго класса системы HLA находятся на поверхности только некоторых клеток иммунной системы, в основном это лейкоциты и макрофаги.
Молекулярная система HLA кодируется в ДНК короткого плеса 6-й хромосомы, где находится информация о белках, предназначенных для распознавания своих и чужеродных антигенов и для координации клеток иммунитета.
Какой биоматериал можно использовать для исследования?
Венозную кровь, щечный эпителий.
Как правильно подготовиться к исследованию?
- Специальной подготовки не требуется.
Общая информация об исследовании
Одним из иммунологических факторов невынашивания беременности является совпадение по 3 и более общим антигенам HLA II класса. Для иммунной системы любые белки, кодируемые генами, являются антигенами и потенциально способны вызвать иммунный ответ. Ребенок получает половину генов от отца и половину от матери. В первом триместре беременности чужеродные для организма матери отцовские антигены плода вызывают у матери выработку защитных, блокирующих антител.
Что делать при иммунологическом бесплодии?
— Перед планируемой беременностью необходимо вылечить инфекционно-воспалительные процессы у супругов, так как наличие инфекций активирует иммунную систему.
— На 5-8 день менструального цикла (первой фазе) за 2-3 месяца до планируемого зачатия проводят лимфоцитоиммунотерапию (ЛИТ) лимфоцитами мужа (подкожно вводят лейкоциты отца будущего ребенка). ЛИТ наиболее эффективна при наличии 4 и более совпадений по системе HLA и повышает шанс успешной беременности в 3-4 раза.
ЛИТ наиболее эффективна при наличии 4 и более совпадений по системе HLA и повышает шанс успешной беременности в 3-4 раза.
— С 16 по 25-й день менструального цикла (вторая фаза) проводят лечение гормоном дидрогестероном.
— На ранних этапах беременности используют методы активной и пассивной иммунизации: лимфоцитоиммунотерапию (проводят каждые 3-4 недели до 12-14 недель беременности) и внутривенное капельное введение средних доз иммуноглобулина. Эти мероприятия способствуют успешному протеканию I триместра беременности и снижают риск формирования плацентарной недостаточности.
Генетические маркеры
Что означают результаты?
Выдается результат по каждому из трех локусов гена HLA и заключение генетика по исследованным маркерам совместимости — выявленные генотипы, насколько благоприятные/неблагоприятные факторы для репродукции определены.
Литература
1. Wen, L., Wong, F. S., Tang, J., Chen, N.-Y., Altieri, M. , David, C., Flavell, R., Sherwin, R. In vivo evidence for the contribution of human histocompatibility leukocyte antigen (HLA)-DQ molecules to the development of diabetes. 2000.
, David, C., Flavell, R., Sherwin, R. In vivo evidence for the contribution of human histocompatibility leukocyte antigen (HLA)-DQ molecules to the development of diabetes. 2000.
2. Stanescu, H. C., Arcos-Burgos, M., Medlar, A., Bockenhauer, D., Kottgen, A., Dragomirescu, L., Voinescu, C., Patel, N., Pearce, K., Hubank, M., Stephens, H. A. F., Laundy, V., and 17 others. Risk HLA-DQA1 and PLA2R1 alleles in idiopathic membranous nephropathy. New Eng. J. Med. 364: 616-626, 2011.
3. Erlich, H., Lee, J. S., Petersen, J. W., Bugawan, T., DeMars, R. Molecular analysis of HLA class I and class II antigen loss mutants reveals a homozygous deletion of the DR, DQ, and part of the DP region: implications for class II gene order. Hum. Immun. 16: 205-219, 1986.
4. Nabozny, G. H., Baisch, J. M., Cheng, S., Cosgrove, D., Griffiths, M. M., Luthra, H. S., David, C. S. HLA-DQ8 transgenic mice are highly susceptible to collagen-induced arthritis: a novel model for human polyarthritis. J. Exp. Med. 183: 27-37, 1996.
Exp. Med. 183: 27-37, 1996.
5. Shackelford, D. A., Mann, D. L., van Rood, J. J., Ferrara, G. B., Strominger, J. L. Human B-cell alloantigens DC1, MT1, and LB12 are identical to each other but distinct from the HLA-DR antigen. Proc. Nat. Acad. Sci. 78: 4566-4570, 1981.
Генетический контроль иммунного ответа — Med24info.com
С помощью технологии отключения (нокаута) генов показано, что для реализации иммунного ответа необходимо функционирование большого числа генов. О том же свидетельствуют данные оценки экспрессии генов при активации клеток иммунной системы — в этом процессе участвуют сотни генов. Генетический контроль в более узком смысле подразумевает возможность варьирования выраженности различных звеньев иммунного ответа в зависимости от аллельных форм тех или иных генов. Наибольшее внимание уделяют двум направлениям изучения генетического контроля иммунных процессов — анализу генетических основ контроля иммунного ответа безотносительно к его специфичности и в зависимости от специфичности вызывающих его антигенов.
 2-зависимых форм иммунного ответа.
2-зависимых форм иммунного ответа.
Значительно полнее разработана вторая линия исследований — изучение генетической детерминации уровня иммунного ответа на конкретные антигенные эпитопы. Главные результаты в этой области получены при изучении ответа мышей и морских свинок различных линий на синтетические пептиды с известной структурой эпитопов. Примером
Рис. 3.133. Уровень гуморального иммунного ответа на синтетические пептиды детерминирован генами MHC (у мышей — Н-2). Структурно сходные пептиды индуцируют гуморальный иммунный ответ при оппозитном по направленности контроле со стороны молекул MHC-II
таких результатов могут служить данные о линейных различиях уровня ответа на два синтетических полипептида — (TG)—A—L и (HG) — A — L (см. раздел 3.2.1.3). Оказалось, что генетический контроль иммунного ответа индивидуален для каждого антигена. Контрастным характером ответа отличаются мыши линий C57BL/6 (H-2b) и CBA (H-2k): первые сильно отвечают на (TG)—A—L и слабо — на (HG)—A—L, вторые (CBA) — наоборот (рис.
 3.133). Уровень иммунного ответа на названные антигены контролирует один ген, первоначально обозначенный как Ir-1 (Immune response 1). Аллель, детерминирующий высокий уровень ответа, является доминантным. Ген Ir-1 оказался сцепленным с генетическим комплексом MHC, точнее локализуется в области генов MHC класса II. Позже была установлена идентичность гена Ir-1 генам H-2I у мышей. Это подтверждают данные о том, что мутация, затрагивающая экзон гена H-2I-A, кодирующий аминокислотную последовательность в вариабельной области молекулы I-A, приводит к смене высокой отвечаемости на низкую.
3.133). Уровень иммунного ответа на названные антигены контролирует один ген, первоначально обозначенный как Ir-1 (Immune response 1). Аллель, детерминирующий высокий уровень ответа, является доминантным. Ген Ir-1 оказался сцепленным с генетическим комплексом MHC, точнее локализуется в области генов MHC класса II. Позже была установлена идентичность гена Ir-1 генам H-2I у мышей. Это подтверждают данные о том, что мутация, затрагивающая экзон гена H-2I-A, кодирующий аминокислотную последовательность в вариабельной области молекулы I-A, приводит к смене высокой отвечаемости на низкую.
Дальнейший анализ показал, что действие гена реализуется на уровне АПК, экспрессирующих молекулы MHC-II. Его механизм состоит в различной способности антигенных пептидов (Т-эпитопов) встраивать
ся в антигенсвязывающую щель в молекуле MHC-II. Хорошее встраивание соответствует высокому уровню отвечаемости, плохое встраивание — низкому. Таким образом, среди многочисленных событий, составляющих иммунный ответ, ключевым для детерминации его количественных показателей является этап презентации антигенного эпитопа Т-хел- перу.
Ir-1/MHC-II — основной, но не единственный локус, определяющий уровень иммунного ответа. Описаны локализованные в других участках MHC гены, определяющие уровень ответа (практически всегда — через влияние на презентацию антигена).
 К генам иммунного ответа относят гены, контролирующие процессинг антигенов, например LMP и ТАР (оба они также расположены в регионе II комплекса MHC). Гены группы LMP кодируют компоненты протеасом, в которых происходит расщепление белков на пептиды, встраивающиеся в молекулы MHC-I, а гены группы ТАР — компоненты транспортной системы, перемещающей эти пептиды из цитозоля в эндоплазматический ретикулум. Регуляция уровня иммунного ответа этими генами обусловлена сродством их продуктов к соответствующим белкам или пептидам и, следовательно, полнотой выполнения ими функций (расщепления, транспорта) и в конечном счете эффективностью доставки пептидов к молекулам MHC-I. Сродство пептидов к антигенсвязывающей щели молекулы MHC-I также служит одним из факторов генетической детерминации уровня ответа, в частности против вирусов (рис. 3.134).
К генам иммунного ответа относят гены, контролирующие процессинг антигенов, например LMP и ТАР (оба они также расположены в регионе II комплекса MHC). Гены группы LMP кодируют компоненты протеасом, в которых происходит расщепление белков на пептиды, встраивающиеся в молекулы MHC-I, а гены группы ТАР — компоненты транспортной системы, перемещающей эти пептиды из цитозоля в эндоплазматический ретикулум. Регуляция уровня иммунного ответа этими генами обусловлена сродством их продуктов к соответствующим белкам или пептидам и, следовательно, полнотой выполнения ими функций (расщепления, транспорта) и в конечном счете эффективностью доставки пептидов к молекулам MHC-I. Сродство пептидов к антигенсвязывающей щели молекулы MHC-I также служит одним из факторов генетической детерминации уровня ответа, в частности против вирусов (рис. 3.134).
 е. развитие иммунного ответа на аутоантигены) четко контролируют названные гены (подробнее см. раздел 4.4.1.2). На этой связи основана проблема «HLA и иммунозависимые болезни». У людей с определенным генотипом величины относительного риска развития аутоиммунных заболеваний (отношение частоты развития заболевания у носителей данного аллеля и у лиц, не имеющих его) бывают повышены (равны 10—20). В отдельных случаях они значительно выше, достигая 208 при анкилозирующем спондилартрите (связь с аллелем HLA- В27 у ориентов). Доказательством реальности этой связи являются результаты переноса (трансфекции) мышам человеческого гена В27, который приводит к развитию у этих мышей патологии, сходной с анкилозирующим спондилартритом. Проявление эффекта гена (пенетрантность) зависит от ряда обстоятельств, в том числе генетического фона: у людей различных
е. развитие иммунного ответа на аутоантигены) четко контролируют названные гены (подробнее см. раздел 4.4.1.2). На этой связи основана проблема «HLA и иммунозависимые болезни». У людей с определенным генотипом величины относительного риска развития аутоиммунных заболеваний (отношение частоты развития заболевания у носителей данного аллеля и у лиц, не имеющих его) бывают повышены (равны 10—20). В отдельных случаях они значительно выше, достигая 208 при анкилозирующем спондилартрите (связь с аллелем HLA- В27 у ориентов). Доказательством реальности этой связи являются результаты переноса (трансфекции) мышам человеческого гена В27, который приводит к развитию у этих мышей патологии, сходной с анкилозирующим спондилартритом. Проявление эффекта гена (пенетрантность) зависит от ряда обстоятельств, в том числе генетического фона: у людей различных
Рис. 3.134. Сродство пептидов вируса гриппа к аллельным вариантам молекул главного комплекса гистосовместимости I класса (MHC-I). В разные аллельные варианты молекулы MHC-I встраиваются различные пептиды белка NP вируса гриппа, распознаваемые CD8+ Т-клетками, что определяет специфичность иммунного ответа
этнических групп величина относительного риска развития заболевания при наличии аллеля В27 варьирует.
 2, в результате чего носители названных аллелей и особенно гаплотипа А2В27 оказываются устойчивыми к гриппу. В то же время преимущественное встраивание в молекулы HLA-B8 и HLA-DR3 аутоантигенных пептидов из белков щитовидной и поджелудочной желез обусловливает наличие положительной корреляции заболеваемости базедовой болезнью, тиреоидитом Хашимото и ювенильным сахарным диабетом с наличием этих аллелей.
2, в результате чего носители названных аллелей и особенно гаплотипа А2В27 оказываются устойчивыми к гриппу. В то же время преимущественное встраивание в молекулы HLA-B8 и HLA-DR3 аутоантигенных пептидов из белков щитовидной и поджелудочной желез обусловливает наличие положительной корреляции заболеваемости базедовой болезнью, тиреоидитом Хашимото и ювенильным сахарным диабетом с наличием этих аллелей.
Показана роль в контроле уровня иммунного ответа локуса IGH, детерминирующего не только структуру константных доменов иммуноглобулинов, но и некоторые перекрестно-реагирующие идиотипы, связанные с синтезом антител определенной специфичности. С этим генетическим локусом связан контроль уровня гуморального ответа на а(1-3)-декстран, а также IgE-ответа на некоторые аллергены. В случаях МНС-сцепленного контроля он не всегда реализуется через гены МНС: так, предрасположенность к аутоиммунному синдрому новозеландских мышей линии BW1 реализуется через ген TNF.
Генетическая детерминация иммунного ответа не ограничивается контролем его уровня.
 2-клеток (STAT6, GATA-3) и ряд других.
2-клеток (STAT6, GATA-3) и ряд других. Генетические факторы развития сахарного диабета
Роль генетики в диагностике заболеваний
В настоящее время, благодаря развитию молекулярных методов исследования, разработано уже около 200 тестов, позволяющих выявлять наследственные предрасположенности к различным заболеваниям. Следует отметить, что исследования, направленные для выявления предрасположенности к развитию заболеваний, не ставят целью поставить диагноз, они лишь указывают на наличие генов, которые ассоциированы с заболеваниями.
«Генетические маркёры» сахарного диабета
Показано, что ряд заболеваний может передаваться по наследству, а у части популяции имеются предпосылки для возникновения того или иного заболевания. Были обнаружены гены и их белковые продукты, которые отвечают за развитие таких заболеваний. В лабораторной практике иногда их называют «генетическими маркерами». Изучение таких маркёров дает возможность выделить группы различного риска развития заболеваний, и в частности, диабета. Такой подход может упростить раннюю диагностику заболевания (риск развития заболевания), до проявления основных клинических признаков.
Такой подход может упростить раннюю диагностику заболевания (риск развития заболевания), до проявления основных клинических признаков.
При помощи генетических маркёров можно выявить группы людей с наличием риска развития сахарного диабета. Это является важным этапом диагностики диабета, поскольку в сочетании с традиционными методами (определение глюкозы, гликированного гемоглобина, и т.д.) приводит к улучшению диагностики заболевания еще до проявления выраженных клинических симптомов заболевания и помогает разработке поведения человека и принятия профилактических мер.
Помимо моногенных наследственных заболеваний, обусловленных мутациями в определенном гене, ответственным за кодирование какого-либо белка, часть заболеваний является мультифакторными или сложно наследуемыми. Это означает, что заболевание может реализоваться в результате деятельности нескольких генов, а также в результате воздействия других причин, например, воздействия окружающей среды. К мультифакторным заболеваниям относят: сахарный диабет, остеопороз, атеросклероз, ишемическую болезнь сердца, злокачественные новообразования. К генетически детерминируемым состояниям можно отнести ожирение, склонность к алкоголизму.
К генетически детерминируемым состояниям можно отнести ожирение, склонность к алкоголизму.
Генетика сахарного диабета 2 типа
Генетическая предрасположенность к сахарному диабету 2 типа носит семейный характер, и часто с сопутствующим ожирением. Уже обнаружено около 20 генов, полиморфизмы в которых являются факторами риска возникновения диабета 2 типа.
В оценке возможности развития сахарного диабета определенную роль имеет место изучение полиморфизмов в системе HLA (human leucocyte antigens). Антигены гистосовместимости (HLA-комплекс) — система человека, состоящая из комплекса генов и их продуктов (белков), выполняющих различные биологические функции, и в первую очередь, обеспечивающих генетический контроль иммунного ответа и взаимодействие между собой клеток, которые реализуют этот ответ.
Молекулярно-биологические приемы диагностики сахарного диабета постоянно совершенствуются и вводятся в клиническую практику. Все полученные данные нужно оценивать с врачом-генетиком, учитывая другие клинические и лабораторные данные.
Источник: medaboutme.ru
| Наименование | Биоматериал | Срок | |
|---|---|---|---|
| ANA-профиль, кровь | кровь | 3 рабочих дня* | Заказать |
| Альфа-1 кислый гликопротеин (орозомукоид), кровь, мг/дл | кровь | 1 рабочий день* | Заказать |
| Альфа-1-антитрипсин, кровь, г/л | кровь | 1 рабочий день* | Заказать |
| Антиген HLA B27 / HLA B7 | кровь | 1 рабочий день* | Заказать |
| Антинуклеарные антитела ANA (АНФ), кровь | сыворотка крови | 10 дней* | Заказать |
| Антиовариальные антитела, кровь, Ед/мл | Заказать | ||
| Антиспермальные антитела на сперматозоидах (сперма) | сперма | 1 рабочий день* | Заказать |
| Антиспермальные антитела, кровь, Е/мл | кровь | 1 рабочий день* | Заказать |
Антиспермальные антитела, семенная плазма, колич. , ИФА , ИФА |
* | Заказать | |
| Антистрептолизин-О (АСЛО) колич., кровь, МЕ/мл | кровь | 1 рабочий день* | Заказать |
| Антитела к MCV, кровь, ед/мл | кровь | 20 рабочих дней* | Заказать |
| Антитела к аннексину V IgG, кровь, ед/мл | 15 рабочих дней* | Заказать | |
| Антитела к аннексину V IgM, кровь, ед/мл | кровь | 15 рабочих дней* | Заказать |
| Антитела к антигенам нейтрофилов спектр, кровь | кровь | 2 рабочих дня* | Заказать |
| Антитела к клеткам островков Лангерганса (бета-клетки), IgG, кровь | кровь | 20 рабочих дней* | Заказать |
| Антитела к протромбину, кровь, ед/мл | кровь | 15 рабочих дней* | Заказать |
| Антитела к тканевой трансглутаминазе, кровь | кровь | 15 рабочих дней* | Заказать |
| Антитела к фосфатидил-инозитолу, IgG, кровь | кровь | 1 рабочий день* | Заказать |
| Антитела к фосфатидил-серину, IgG, кровь | кровь | 1 рабочий день* | Заказать |
| Антитела к фосфатидиловой кислоте, IgG, кровь | кровь | 1 рабочий день* | Заказать |
| Антитела к фосфолипидам скрининг (5 антигенов), кровь | кровь | 1 сутки* | Заказать |
| АТ к бета2-гликопротеину 1 суммарные, кровь, Ед/мл | кровь | 20 рабочих дней* | Заказать |
| АТ к ДНК, кровь | сыворотка крови | 10 рабочих дней* | Заказать |
| Ат к инсулину, кровь, Ед/мл | кровь | 1 рабочий день* | Заказать |
| АТ к кардиолипину, суммарные | кровь | 15 рабочих дней. * * |
Заказать |
| АТ к микросомам печени и почек (LKM-1), IgG, кровь | кровь | 15 рабочих дней* | Заказать |
| АТ к митохондриям (AMA M2), IgG, кровь | кровь | 15 рабочих дней* | Заказать |
| Ат к фосфолипидам сумм. (кардиолипин, фосфатидилсерин, фосфатидилинозитол, фосфатидиловая к-та)кровь | кровь | 1 сутки* | Заказать |
| АТ к циклическому цитруллин-содержащему пептиду, IgG, кровь, Е/мл | кровь | 5 рабочих дней* | Заказать |
| АТ к цитоплазматическим антигенам нейтрофилов | кровь | 15 рабочих дней* | Заказать |
| Гаптоглобин, кровь, мг/дл | кровь | 1 рабочий день* | Заказать |
| Гастро-5-лайн | кровь | 1 рабочий день* | Заказать |
| Печеночный аутоиммунный профиль (9 параметров), кровь | кровь | 1 рабочий день* | Заказать |
| Ревматоидный фактор, количественно, МЕ/мл | кровь | 1 день* | Заказать |
| С-реактивный белок суперчувствительный (Siemens), кровь, мг/л | кровь | 1 рабочий день* | Заказать |
| С-реактивный белок, кровь, мг/л | кровь | 1 рабочий день* | Заказать |
| титрование (1 разведение) | * | Заказать | |
| Церулоплазмин, кровь, мг/дл | кровь | 1 рабочий день* | Заказать |
| Наименование | Биоматериал | Срок | |
| АЛАТ, кровь, Е/л | кровь | 1 рабочий день* | Заказать |
| Альбумин, кровь, г/л | кровь | 1 рабочий день* | Заказать |
| Амилаза (диастаза), моча, Ед/л | * | Заказать | |
| Амилаза панкреатическая, кровь, Е/л | кровь | 1 рабочий день* | Заказать |
| Амилаза, кровь, Е/л | * | Заказать | |
| Амилаза-альфа, кровь, Е/л | кровь | 1 рабочий день* | Заказать |
| Аполипопротеины А1 и В | кровь | 1 рабочий день* | Заказать |
| АСАТ, кровь, Е/л | кровь | 1 рабочий день* | Заказать |
АЧТВ, сек.
|
кровь | 1 рабочий день* | Заказать |
| Бикарбонаты, кровь, мМоль/л | кровь, сыворотка | 1 день* | Заказать |
| Билирубин общий, кровь, мкМоль/л | кровь | 1 рабочий день* | Заказать |
| Билирубин свободный, кровь, мкМоль/л | кровь | 1 рабочий день* | Заказать |
| Билирубин связанный, кровь, мкМоль/л | кровь | 1 рабочий день* | Заказать |
| Гастрин, кровь, мЕД/мл | кровь | 1 рабочий день* | Заказать |
| ГГТП, кровь, Е/л | кровь | 1 рабочий день* | Заказать |
| Глюкоза, кровь, мМоль/л | кровь | 1 рабочий день* | Заказать |
| Креатинин, кровь, мкМоль/л | кровь | 1 рабочий день* | Заказать |
| Креатинкиназа, кровь, Е/л | кровь | 1 рабочий день* | Заказать |
| КФ (кислая фосфатаза) общая, кровь, Е/л | кровь | 1 рабочий день* | Заказать |
| КФК-МВ, ЕД/л | Заказать | ||
| Лактат (молочная кислота), кровь, Ед/л | кровь | 1 рабочий день* | Заказать |
| ЛДГ, кровь, Е/л | кровь | 1 рабочий день* | Заказать |
| Липаза панкреатическая, кровь, Е/л | кровь | 1 рабочий день* | Заказать |
| Мочевая кислота, кровь, мкМоль/л | кровь | 1 рабочий день* | Заказать |
| Мочевина(Diasys), кровь, мМоль/л | кровь | 1 рабочий день* | Заказать |
| Мочевина, кровь, мМоль/л | в день обращения, в течении часа* | Заказать | |
| Насыщение трансферрина железом (%) | * | Заказать | |
| Общий белок, кровь, г/л | кровь | 1 рабочий день* | Заказать |
| Протромбин, МНО | кровь | 1 рабочий день* | Заказать |
Тимоловая проба, кровь, ед.
|
кровь | 1 рабочий день* | Заказать |
| Трансферрин, кровь, мг/дл | кровь | 1 рабочий день* | Заказать |
| Триглицериды, кровь, мМоль/л | кровь | 1 рабочий день* | Заказать |
| Тромбиновое время, сек | кровь | 1 рабочий день* | Заказать |
| Ферритин, кровь, мкг/л | кровь | 1 рабочий день* | Заказать |
| Фибриноген, кровь, г/л | кровь | 1 рабочий день* | Заказать |
| Холестерин ЛПВП (HDL), кровь, мМоль/л | кровь | 1 рабочий день* | Заказать |
| Холестерин ЛПНП (LDL), кровь, мМоль/л | кровь | 1 рабочий день* | Заказать |
| Холестерин общий, кровь, мМоль/л | кровь | 1 рабочий день* | Заказать |
| Холестериновый коэффициент атерогенности | кровь | 1 рабочий день* | Заказать |
| Щелочная фосфатаза, кровь, Е/л | кровь | 1 рабочий день* | Заказать |
| Наименование | Биоматериал | Срок | |
| D1 sIgE к аллергенам Dermatophagoides pteronissinus, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| D2 sIgE к аллергенам Dermatophagoides farinae, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| E1 sIgE к аллергенам эпителия кошки, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| E2 sIgE к аллергенам эпителия собаки, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| E5 sIgE к аллергенам перхоти собаки, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| E6 sIgE к аллергенам морской свинки, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| E78 sIgE к аллергенам волнистого попугая, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| E81 sIgE к аллергенам эпителия овцы, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| E82 sIgE к аллергенам эпителия кролика, кровь, kU/L | * | Заказать | |
| E84 sIgE к аллергенам хомяка, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| E85 sIgE к аллергенам пера курицы, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F1 sIgE к аллергенам яичного белка, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F10 sIgE к аллергенам кунжута (сезама), кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F105 sIgE к аллергенам шоколада, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F11 sIgE к аллергенам гречневой муки, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F12 sIgE к аллергенам гороха, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F13 sIgE к аллергенам арахиса, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F14 sIgE к аллергенам соевых бобов, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F17 sIgE к аллергенам фундука, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F2 sIgE к аллергенам коровьего молока, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F20 sIgE к аллергенам миндаля, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F204 sIgE к аллергенам форели, кровь, kU/L | * | Заказать | |
| F221 sIgE к аллергенам кофе, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F225 sIgE к аллергенам тыквы, кровь, kU/L | * | Заказать | |
| F237 sIgE к аллергенам абрикоса, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F25 sIgE к аллергенам помидоров, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F256 sIgE к аллергенам грецкого ореха, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F26 sIgE к аллергенам свинины, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F260 sIgE к аллергенам брокколи, кровь, kU/L | * | Заказать | |
| F27 sIgE к аллергенам говядины, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F284 sIgE к аллергенам индейки, кровь, kU/L | * | Заказать | |
| F3 sIgE к аллергенам трески, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F31 sIgE к аллергенам моркови, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F33 sIgE к аллергенам апельсина, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F35 sIgE к аллергенам картофеля, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F4 sIgE к аллергенам пшеничной муки, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F40 sIgE к аллергенам тунца, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F49 sIgE к аллергенам яблока, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F5 sIgE к аллергенам ржаной муки, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F61 sIgE к аллергенам сардины, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F7 sIgE к аллергенам овсяной муки, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F75 sIgE к аллергенам яичного желтка, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F78 sIgE к аллергенам казеина, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F8 sIgE к аллергенам кукурузной муки, кровь, kU/L | * | Заказать | |
| F83 sIgE к аллергенам куриного мяса, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F87 sIgE к аллергенам дыни, кровь, kU/L | * | Заказать | |
| F88 sIgE к аллергенам баранины, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F9 sIgE к аллергенам риса, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F92 sIgE к аллергенам бананов, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F93 sIgE к аллергенам какао, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F94 sIgE к аллергенам груши, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| F95 sIgE к аллергенам персика, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| FP5 sIgE к миксту пищевых аллергенов (F1, F2, F3, F4, F13, F14), кровь | кровь | 1 рабочий день* | Заказать |
| FP73 sIgE к миксту пищевых аллергенов (F26, F27, F83, F88), кровь | кровь | 1 рабочий день* | Заказать |
| G12 sIgE к аллергенам ржи культивированной, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| G15 sIgE к аллергенам пшеницы, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| G3 sIgE к аллергенам ежи сборной, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| G4 sIgE к аллергенам овсяницы луговой, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| G5 sIgE к аллергенам ржи многолетней, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| G6 sIgE к аллергенам тимофеевки, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| G8 sIgE к аллергенам мятлика лугового, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| GP1 sIgE к миксту аллергенов травы (G3, G4, G5, G6, G8), кровь | кровь | 1 рабочий день* | Заказать |
| h2 sIgE к аллергенам домашней пыли (Greer), кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| h3 sIgE к аллергенам домашней пыли (Hollister-Stier), кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| HP1 sIgE к миксту аллергенов пыли (h2, D1, D2, I6), кровь | кровь | 1 рабочий день* | Заказать |
| I6 sIgE к аллергенам таракана-прусака, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| IP8 sIgE к миксту ингаляционных аллергенов (E1, D1, T3, E5, W6, G6, G12, M2), кровь | кровь | 1 рабочий день* | Заказать |
| M1 sIgE к аллергенам Penicillium notatum, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| M2 sIgE к аллергенам Cladosporium herbarum, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| M3 sIgE к аллергенам Aspergillus fumigatus, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| M5 sIgE к аллергенам Candida albicans, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| M6 sIgE к аллергенам Alternaria tenuis, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| M70 sIgE к аллергенам Pityrosporum orbiculare, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| MP1 sIgE к миксту аллергенов микроскопических грибов (M1, M2, M3, M5, M6), кровь | кровь | 1 рабочий день* | Заказать |
| O1 sIgE к аллергенам хлопка, кровь, kU/L | * | Заказать | |
O72 sIgE к энтеротоксину А Staph. aureus, кровь, kU/L aureus, кровь, kU/L
|
кровь | 1 рабочий день* | Заказать |
| O73 sIgE к энтеротоксину B Staph. aureus, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| P1 sIgE к аллергенам аскариды, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| T1 sIgE к аллергенам клена ясенелистного, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| T12 sIgE к аллергенам ивы, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| T14 sIgE к аллергенам тополя, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| T19 sIgE к аллергенам акации, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| T2 sIgE к аллергенам ольхи, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| T3 sIgE к аллергенам березы, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| T4 sIgE к аллергенам лещины обыкновенной, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| T7 sIgE к аллергенам дуба, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| TP9 sIgE к миксту аллергенов деревьев (T2, T12, T4, T7, T3), кровь | кровь | 1 рабочий день* | Заказать |
| W1 sIgE к аллергенам амброзии обыкновенной, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| W10 sIgE к аллергенам мари белой, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| W11 sIgE к аллергенам солянки/зольника, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| W15 sIgE к аллергенам лебеды, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| W6 sIgE к аллергенам полыни обыкновенной, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| W8 sIgE к аллергенам одуванчика лекарственного, кровь, kU/L | * | Заказать | |
| W9 sIgE к аллергенам подорожника, кровь, kU/L | кровь | 1 рабочий день* | Заказать |
| WP1 sIgE к миксту аллергенов сорных трав (W1, W11, W8, W9, W6), кровь | кровь | 1 рабочий день* | Заказать |
| К20 sIgE к аллергенам шерсти, кровь, kU/L | * | Заказать | |
| К74 sIgE к аллергенам шелка, кровь, kU/L | * | Заказать | |
| К82 sIgE к аллергенам латекса, кровь, kU/L | * | Заказать | |
| Общий IgE (Siemens), кровь, МЕ/мл | кровь | 1 рабочий день* | Заказать |
| Пищевая IgG аллергия, 90 аллергенов (Российская панель) | кровь | 2 рабочих дня* | Заказать |
| С1-ингибитор эстеразы, кровь | кровь | 1 рабочий день* | Заказать |
| Эозинофильный катионный белок (Siemens), кровь, нг/мл | кровь | 1 рабочий день* | Заказать |
| Наименование | Биоматериал | Срок | |
| HbeAg, кровь | кровь | 5 рабочих дней* | Заказать |
| HbsAg, Cito!, кровь | * | Заказать | |
| HbsAg, кровь | кровь | 5 рабочих дней* | Заказать |
| АТ Chlamydia pneumonia IgG, кровь, Е/мл | кровь | 10 рабочих дней* | Заказать |
| АТ Chlamydia pneumonia IgА, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ Chlamydia pneumonia IgМ, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ Chlamydia trachomatis IgA, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ Chlamydia trachomatis IgG cHSP60, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ Chlamydia trachomatis IgG Cito, кровь | * | Заказать | |
| АТ Chlamydia trachomatis IgG Ед/Мл (количественно), кровь | кровь | 10 рабочих дней* | Заказать |
| АТ Chlamydia trachomatis IgG МОМР+pgp3, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ Chlamydia trachomatis IgG, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ Chlamydia trachomatis IgM, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ Herpes simplex IgG (титр), кровь | кровь | 10 рабочих дней* | Заказать |
| АТ Herpes simplex IgG авидность, кровь, % | кровь | 10 рабочих дней* | Заказать |
| АТ Herpes simplex IgM, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ Herpes simplex II типа IgG, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ Herpes simplex II типа IgM, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ Mycoplasma hominis (IgA, IgG) | кровь | 10 рабочих дней* | Заказать |
| АТ Mycoplasma hominis IgM, кровь | Заказать | ||
| АТ Mycoplasma pneumonia IgA, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ Mycoplasma pneumonia IgM, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ Mycoplasma pneumoniae IgG, количественно, МЕ/мл, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ Toxoplasma gondii IgG (Siemens), кровь, МЕ/мл | кровь | 1 рабочий день* | Заказать |
| АТ Toxoplasma gondii IgG авидность, кровь, % | кровь | 10 рабочих дней* | Заказать |
| АТ Toxoplasma gondii IgG+IgM | кровь | 10 рабочих дней* | Заказать |
| АТ Toxoplasma gondii IgM (Siemens), кровь | кровь | 1 рабочий день* | Заказать |
АТ Toxoplasma gondii IgА (кач. ), кровь ), кровь
|
кровь | 10 рабочих дней* | Заказать |
| АТ Treponema pallidum Cito!, суммарные | * | Заказать | |
| АТ Trichomonas vaginalis IgG, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ Trichomonas vaginalis IgА, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ Ureaplasma urealyticum | кровь | 10 рабочих дней* | Заказать |
| АТ Varicella zoster virus (VZV) IgG, кровь | кровь | 5 рабочих дней* | Заказать |
| АТ Varicella zoster virus (VZV) IgM, кровь | кровь | 5 рабочих дней* | Заказать |
| АТ Вирус Эпштейна-Барр (капсидный антиген) IgM, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ Вирус Эпштейна-Барр (ранний антиген EA) IgG, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ Вирус Эпштейна-Барр (ядерный антиген NA) IgG, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ к HBcAg IgG, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ к HBcAg IgM, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ к HBcAg суммарные, кровь | кровь | 10 рабочих дней* | Заказать |
АТ к B. pertussis (коклюш) IgA (кач.), кровь pertussis (коклюш) IgA (кач.), кровь
|
кровь | 14 рабочих дней* | Заказать |
| АТ к B. pertussis (коклюш) IgG Ед/мл, кровь | кровь | 14 рабочих дней* | Заказать |
| АТ к B. pertussis (коклюш) IgM, (кач.), кровь | кровь | 14 рабочих дней* | Заказать |
| АТ к Borrelia burgdorferi s.l., IgG, кровь | кровь | 1 рабочий день* | Заказать |
| АТ к Candida, IgA, кровь | * | Заказать | |
| АТ к Candida, IgG, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ к Candida, IgM, кровь | * | Заказать | |
АТ к H. pylori IgG колич. (Siemens), кровь pylori IgG колич. (Siemens), кровь
|
кровь | 1 рабочий день* | Заказать |
| АТ к HAV IgG, Cito!, кровь | кровь | 1 рабочий день* | Заказать |
| АТ к HAV IgM, Cito!, кровь | кровь | 1 рабочий день* | Заказать |
| АТ к HbeAg, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ к HbsAg качественно, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ к HbsAg количественно, кровь, МЕ/мл | * | Заказать | |
| АТ к HCV IgG авидность, кровь, % | кровь | 10 рабочих дней* | Заказать |
| АТ к HCV IgM, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ к HCV подтверждающий тест | кровь | 10 рабочих дней* | Заказать |
| АТ к HCV суммарные (скрининг!), кровь | кровь | 10 рабочих дней* | Заказать |
| АТ к HCV суммарные Cito!, кровь | кровь | 10 рабочих дней* | Заказать |
АТ к HDV сумм. , кровь , кровь
|
* | Заказать | |
| АТ к HDV суммарные, Cito!, кровь | кровь | 10 рабочих дней* | Заказать |
| Ат к HEV, IgG, кровь | кровь | 10 рабочих дней* | Заказать |
| Ат к HEV, IgM, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ к HGV IgG, скрининг, кровь | кровь | 15 рабочих дней* | Заказать |
| АТ к Mycobacterium tuberculosis Cito!, кровь | кровь | 1 рабочий день* | Заказать |
| АТ к Treponema pallidum IgG, качественно | Заказать | ||
| АТ к Treponema pallidum, суммарные, качественно | кровь | 10 рабочих дней* | Заказать |
| АТ к Yersinia enterocolitica (факторы вирулентности) IgG Ед/Мл (количественно), кровь | кровь | 15 рабочих дней* | Заказать |
| Ат к Yersinia Enterocolitica + Pseudotuberculosis IgA, кровь | * | Заказать | |
| Ат к Yersinia Enterocolitica + Pseudotuberculosis IgG, кровь | * | Заказать | |
| Ат к Yersinia Enterocolitica + Pseudotuberculosis IgM, кровь | * | Заказать | |
| АТ к антигенам HCV | * | Заказать | |
| АТ к аскариде IgG скрининг, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ к аскариде IgG титр, кровь | кровь | 10 рабочих дней* | Заказать |
АТ к аспергилле сумм. титр, кровь титр, кровь |
* | Заказать | |
| АТ к аспергилле, IgG, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ к вирусу кори IgG (количественно) | кровь | 9 рабочих дней* | Заказать |
| АТ к вирусу кори IgM (качественно) | * | Заказать | |
| Ат к вирусу паротита, IgG, кровь | кровь | 9 рабочих дней* | Заказать |
| Ат к вирусу паротита, IgM, кровь | кровь | 9 рабочих дней* | Заказать |
| АТ к вирусу ТТ (торкутенувирус гепатита неясной этиологии) IgG, кровь | * | Заказать | |
| АТ к ВИЧ, кровь, cito | * | Заказать | |
| Ат к возбудителю бруцеллеза IgA, кровь | * | Заказать | |
| Ат к возбудителю бруцеллеза IgG, кровь | кровь | 11 рабочих дней* | Заказать |
| Ат к возбудителю бруцеллеза IgM, кровь | * | Заказать | |
| АТ к гельминтам | кровь | 10 рабочих дней* | Заказать |
| АТ к глиадину | кровь | 10 рабочих дней* | Заказать |
| АТ к краснухе IgG (Siemens), кровь, МЕ/мл | кровь | 1 рабочий день* | Заказать |
| АТ к краснухе IgG авидность, кровь, % | кровь | 1 рабочий день* | Заказать |
| АТ к краснухе IgM (ИФА), кровь | Заказать | ||
| АТ к краснухе IgM, кровь | кровь | 1 рабочий день* | Заказать |
| АТ к лямблиям IgM, кровь | кровь | 10 рабочих дней* | Заказать |
АТ к лямблиям сумм. скрининг, кровь скрининг, кровь
|
кровь | 10 рабочих дней* | Заказать |
| АТ к лямблиям сумм. титр, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ к Описторхису IgG скрининг, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ к Описторхису IgG титр, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ к Сlonorchis sinensis IgG (китайской двуустке) скрининг, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ к Сlonorchis sinensis IgG (китайской двуустке) титр, кровь | кровь | 10 рабочих дней* | Заказать |
| Ат к сальмонеллам, кровь | кровь | 11 рабочих дней* | Заказать |
| АТ к токсокарам IgG скрининг, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ к токсокарам IgG титр, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ к трихинелле IgG, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ к трихинелле IgG, титр, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ к трихинелле IgМ, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ к эхинококку, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ к эхинококку, титр, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ ЦМВ IEA (предранний белок) | кровь | 10 рабочих дней* | Заказать |
| АТ ЦМВ IgG (титр), кровь | кровь | 10 рабочих дней* | Заказать |
| АТ ЦМВ IgG авидность, кровь, % | кровь | 10 рабочих дней* | Заказать |
| АТ ЦМВ IgG количественно, кровь, РЕ/мл | кровь | 10 рабочих дней* | Заказать |
| АТ ЦМВ IgG, кровь | кровь | 10 рабочих дней* | Заказать |
| АТ ЦМВ IgM, кровь | кровь | 10 рабочих дней* | Заказать |
| Наименование | Биоматериал | Срок | |
| Биоценоз | * | Заказать | |
Биоценоз (G. vag., Atop.vag., Lactobact., бакмасса) количественно vag., Atop.vag., Lactobact., бакмасса) количественно
|
соскоб | 4 рабочих дня* | Заказать |
| Генетика Метаболизма Лактозы | кровь | 5 рабочих дней* | Заказать |
| Генетика Метаболизма Фолатов | кровь | 5 рабочих дней* | Заказать |
| Генотипирование HCV, кровь | кровь | 10 рабочих дней* | Заказать |
| ДНК Atopobium vagine, соскоб (PCR real time) | * | Заказать | |
| ДНК C.alb./C.glabr./C.crusei качественно | соскоб | 1 рабочий день* | Заказать |
| ДНК Candida (C.albicans, C.glabrata, C.krusei, C.parapsilosis и C.tropicalis) количественно, соскоб | * | Заказать | |
| ДНК Candida albicans, кровь | кровь | 1 рабочий день* | Заказать |
| ДНК Candida albicans, соскоб | соскоб | 1 рабочий день* | Заказать |
ДНК Chl. trach./N.gonorr./M.genit./Tr.vag. trach./N.gonorr./M.genit./Tr.vag.
|
соскоб | 1 рабочий день* | Заказать |
| ДНК Chlamydia trachomatis, количественно, соскоб (PCR real time) | * | Заказать | |
| ДНК Chlamydia trachomatis, соскоб (PCR real time) | соскоб | 1 рабочий день* | Заказать |
| ДНК Cytomegalovirus (CMV), кровь (PCR real time) | кровь | 5 рабочих дней* | Заказать |
| ДНК Cytomegalovirus (CMV), ликвор (PCR real time) | ликвор | 5 рабочих дней* | Заказать |
| ДНК Cytomegalovirus (CMV), соскоб (PCR real time) | соскоб | 1 рабочий день* | Заказать |
| ДНК Gardn. vaginalis / Lactobacillus spp. количественно, соскоб (PCR real time) | соскоб | 1 рабочий день* | Заказать |
| ДНК Gardnerella vaginalis, соскоб | соскоб | 1 рабочий день* | Заказать |
ДНК H. pylori, биоптат слизистой оболочки желудка pylori, биоптат слизистой оболочки желудка |
* | Заказать | |
| ДНК H.pylori, кал | кал | 1 рабочий день* | Заказать |
| ДНК HBV (вирус гепатита B), кровь (PCR real time) | кровь | 5 рабочих дней* | Заказать |
| ДНК HBV, копий/мл (PCR real time) | кровь | 15 рабочих дней* | Заказать |
| ДНК Herpes simplex I, II, кровь | кровь | 5 рабочих дней* | Заказать |
| ДНК Herpes simplex I, II, ликвор | ликвор | 5 рабочих дней* | Заказать |
| ДНК Herpes simplex I, II, соскоб | соскоб | 1 рабочий день* | Заказать |
| ДНК Herpes simplex I, II, типирование, кровь | * | Заказать | |
| ДНК Herpes simplex I, II, типирование, соскоб | * | Заказать | |
| ДНК HPV 16/18 количественно, соскоб (PCR real time) | соскоб | 1 рабочий день* | Заказать |
| ДНК HPV 16/18 количественно, соскоб (PCR real time) | * | Заказать | |
| ДНК HPV 6, 11, соскоб | соскоб | 3 рабочих дня* | Заказать |
| ДНК HPV ВКР скрининг количественно, соскоб | соскоб | 3 рабочих дня* | Заказать |
| ДНК HPV высокого онкогенного риска (тип 16,18,31,33,35,39,45,51,52,56,58,59) типирование, соскоб | соскоб | 3 рабочих дня* | Заказать |
| ДНК HPV высокого онкогенного риска тип 16, соскоб | соскоб | 3 рабочих дня* | Заказать |
| ДНК HPV высокого онкогенного риска тип 18, соскоб | соскоб | 3 рабочих дня* | Заказать |
| ДНК HPV скрининг высокого онкогенного риска (тип 16,18,31,33,35,39,45,52,58,59,67), соскоб | соскоб | 3 рабочих дня* | Заказать |
| ДНК Mycoplasma genitalium, количественно, соскоб (PCR real time) | * | Заказать | |
| ДНК Mycoplasma genitalium, соскоб (PCR real time) | соскоб | 1 рабочий день* | Заказать |
| ДНК Mycoplasma hominis, количественно, соскоб (PCR real time) | * | Заказать | |
| ДНК Mycoplasma hominis, соскоб (PCR real time) | соскоб | 1 рабочий день* | Заказать |
| ДНК Mycoplasma pneumoniae/Chlamydophila pneumoniae, мокрота (PCR real time) | Заказать | ||
| ДНК Neisseria gonorrhoeae, соскоб (PCR real time) | соскоб | 1 рабочий день* | Заказать |
| ДНК Trichomonas vaginalis, соскоб (PCR real time) | соскоб | 1 рабочий день* | Заказать |
ДНК Ur. parv./Ur.ur./M.hom. количественно parv./Ur.ur./M.hom. количественно
|
соскоб | 1 рабочий день* | Заказать |
| ДНК Ureaplasma parvum, соскоб (PCR real time) | соскоб | 1 рабочий день* | Заказать |
| ДНК Ureaplasma spp. количественно, соскоб (PCR real time) | соскоб | 1 рабочий день* | Заказать |
| ДНК Ureaplasma urealyticum (T960) + Ureaplasma parvum, соскоб (PCR real time) | соскоб | 1 рабочий день* | Заказать |
| ДНК Ureaplasma urealyticum (T960), соскоб (PCR real time) | соскоб | 1 рабочий день* | Заказать |
| ДНК вируса Varicella zoster (VZV), кровь | кровь | 5 рабочих дней* | Заказать |
| ДНК вируса Varicella zoster (VZV), ликвор | ликвор | 5 рабочих дней* | Заказать |
| ДНК вируса Varicella zoster (VZV), соскоб | * | Заказать | |
| ДНК вируса герпеса тип VI, кровь | кровь | 1 рабочий день* | Заказать |
| ДНК вируса герпеса тип VI, соскоб | соскоб | 1 рабочий день* | Заказать |
| ДНК вируса Эпштейна-Барр количественно, кровь (PCR real time) | кровь | 1 рабочий день* | Заказать |
| ДНК вируса Эпштейна-Барра (EBV), кровь | кровь | 1 рабочий день* | Заказать |
| ДНК вируса Эпштейна-Барра (EBV), соскоб | соскоб | 1 рабочий день* | Заказать |
| ДНК коклюша, паракоклюша и бронхисептикоза, мокрота (PCR real time) | Заказать | ||
| КардиоГенетика Гипертония | кровь | * | Заказать |
| КардиоГенетика Тромбофилия | кровь | 5 дней* | Заказать |
| Полиморфизм гена BRCA, PCR real-time, кровь | кровь | 2 рабочих дня. * * |
Заказать |
| Полиморфизм гена IL28B, PCR real-time, кровь | * | Заказать | |
| РНК HCV, кровь, копий/мл | кровь | 15 рабочих дней* | Заказать |
| РНК HDV, кровь | кровь | 5 рабочих дней* | Заказать |
| РНК HGV, кровь | кровь | 5 рабочих дней* | Заказать |
| РНК HСV (вирус гепатита С), кровь | кровь | 5 рабочих дней* | Заказать |
| УБП-8 | мазок | 7 рабочих дней* | Заказать |
| ФармакоГенетика Варфарин | * | Заказать | |
Фемофлор 4(G.vag., Candida spp.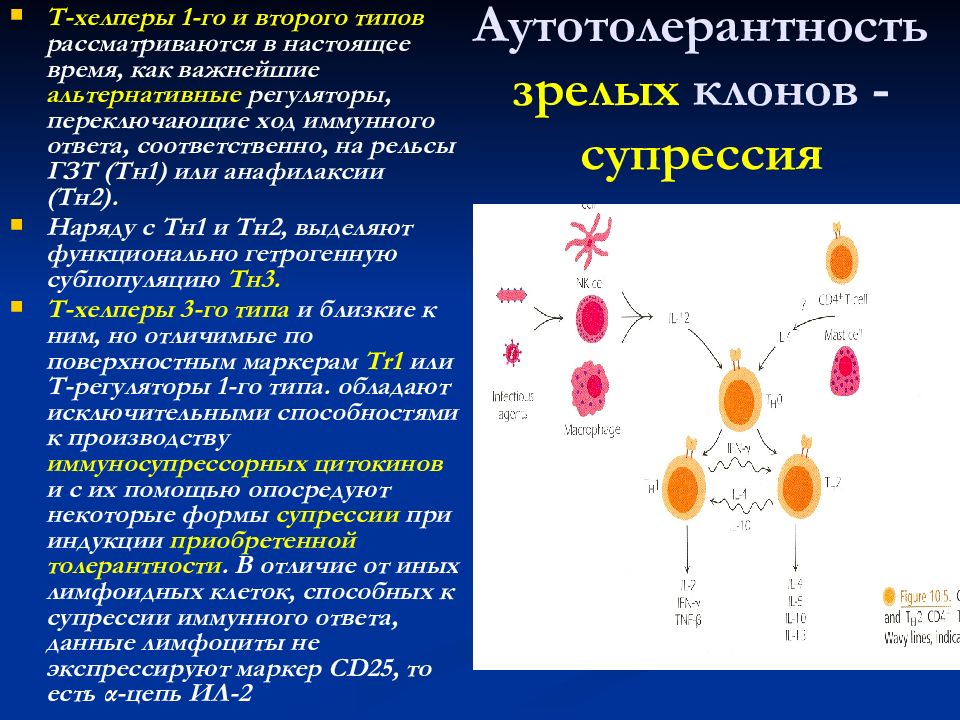 , Lactobact., бакмасса) количественно , Lactobact., бакмасса) количественно |
* | Заказать | |
| Фемофлор 8(gard vag, cand sp, lactob, strept., myc hom., eubact, enterobac, б/масса), количественно | * | Заказать | |
| Наименование | Биоматериал | Срок | |
| Аденовирус респираторный (Adenovirus Ag), ИХА-скрининг, мазок, аспират | * | Заказать | |
| Аллоиммунные антитела (включая антитела к Rh-антигену) | Заказать | ||
| БАК исследование кала на возбудителя Дизентерии (Shigella spp.) | Заказать | ||
| Бак. посев | отделяемое влагалища, уретры, цервикального канала, стенки глотки и любая другая биологическая среда. |
10 рабочих дней* | Заказать |
| Бакпосев мокроты | * | Заказать | |
| Бакпосев мочи с определением чувствительности к антибиотикам | моча | 10 рабочих дней* | Заказать |
| Бакпосев на стафилококк | * | Заказать | |
| Бакпосев отделяемого зева | * | Заказать | |
| Бакпосев отделяемого урогенитального тракта с определением чувствительности к антибиотикам | отделяемое урогенитального тракта | 10 рабочих дней* | Заказать |
| Гомоцистеин, кровь, мкмоль/л | кровь | 13 рабочих дней* | Заказать |
| Гриппа вирус А/В (Influenza Ag А/В), ИХА-скрининг, мазок, аспират | * | Заказать | |
| Грудное молоко на стерильность — одна проба | Грудное молоко | 10 рабочих дней* | Заказать |
| Группа крови и резус-фактор (метод колоночной агглютинаций) | Заказать | ||
| Забор крови на дому I | Заказать | ||
| Забор крови на дому II | Заказать | ||
| Забор крови с иглой бабочкой | Заказать | ||
Забор материала (в т. ч. контейнер с транспортной средой) ч. контейнер с транспортной средой) |
* | Заказать | |
| Забор материала (Исполнитель ГКБ №7) | Заказать | ||
| Забор материала на бакпосев | * | Заказать | |
| Исследование на кандидамикоз | отделяемое влагалища, уретры, цервикального канала, стенки глотки и любая другая биологическая среда. | 10 рабочих дней* | Заказать |
| Кал на дисбактериоз | кал | 10 рабочих дней* | Заказать |
| Кал на кишечные инфекции | кал | 10 рабочих дней* | Заказать |
| Кал на скрытую кровь (Иммунохроматография, Германия) | кал | 5 рабочих дней* | Заказать |
| Кал на яйца гельминтов | кал | 2 рабочих дня* | Заказать |
| Кальпротектин (количественно), кал, мкг/г | кал | 1 рабочий день* | Заказать |
| Копрограмма | кал | 5 рабочих дней* | Заказать |
| Лимонная кислота, фруктоза | * | Заказать | |
| Мазок на онкоцитологию | мазок из шейки матки | 5-7 рабочих дней* | Заказать |
| Мазок на флору | мазок из влагалища у женщин, из уретры – у мужчин.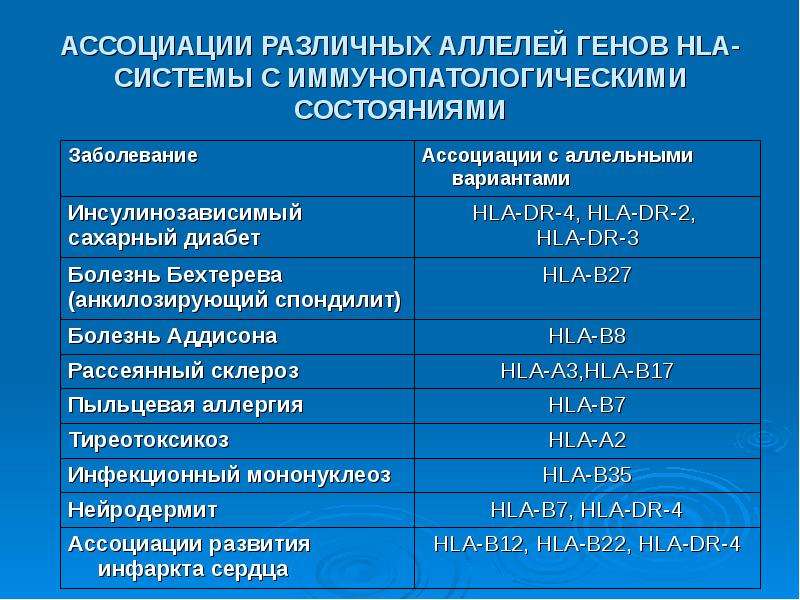 |
2 рабочих дня* | Заказать |
| Мазок на флору гинекологический | Заказать | ||
| Мазок на флору урологический | Заказать | ||
| Микроскопия секрета простаты | * | Заказать | |
| Моча на степень бактериурии | моча | 10 рабочих дней* | Заказать |
| Посев мокроты | * | Заказать | |
| Посев на дисбактериоз | * | Заказать | |
| Посев на уреамикоплазму с определением чувствительности к антибиотикам | * | Заказать | |
| Респираторно-синцитиальный вирус (RSA), ИХА-скрининг, мазок, аспират | * | Заказать | |
| Скрининг 6 наркотических веществ и метаболитов, моча | * | Заказать | |
| Соскоб с языка на кандиды (микроскопия) | соскоб с языка | 10 рабочих дней* | Заказать |
| Стрептококк А (Strep-A Ag), ИХА-скрининг, мазок, аспират | * | Заказать | |
| Типирование HLA II класса локус DQA1 | кровь | 14 рабочих дней* | Заказать |
| Типирование HLA II класса локус DQB1 | кровь | 14 рабочих дней* | Заказать |
| Типирование HLA II класса локус DRB1 | кровь | 14 рабочих дней* | Заказать |
| ФНО (фактор некроза опухоли), кровь, пг/мл | кровь | 10 рабочих дней* | Заказать |
| Цитологическое исследование материала ТАБ | аспират содержимого узловых образований щитовидной железы | 2(рабочих) дня * | Заказать |
| Чувствительность к антибиотикам | любой биологический материал (отделяемое влагалища, уретры, цервикального канала, стенки глотки и т. д.). д.). |
10 рабочих дней* | Заказать |
| Чувствительность к антибиотикам (моча) | * | Заказать | |
| Чувствительность к антимикотикам | любой биологический материал (отделяемое влагалища, уретры, цервикального канала, стенки глотки и т.д.). | 10 рабочих дней* | Заказать |
| Чувствительность к антисептикам | * | Заказать | |
| Чувствительность к бактериофагам | * | Заказать | |
| Эластаза-1 панкреатическая (количественно), кал, мкг Э/г | кал | 14 рабочих дней* | Заказать |
| Эритропоэтин, кровь, мМЕ/мл | кровь | 9 рабочих дней* | Заказать |
| Наименование | Биоматериал | Срок | |
| Выдача повторного снимка | * | Заказать | |
| Дополнительное контрастирование | * | Заказать | |
| МРТ артерий головного мозга | в день обращения, в течении часа* | Заказать | |
| МРТ артерий шеи | в день обращения, в течении часа* | Заказать | |
| МРТ брюшной полости и холангиография | в день обращения, в течении часа* | Заказать | |
| МРТ вен головного мозга | в день обращения, в течении часа* | Заказать | |
| МРТ гипофиза | в день обращения, в течении часа* | Заказать | |
| МРТ глазных орбит и зрительного нерва | в день обращения, в течении часа* | Заказать | |
| МРТ головного мозга | в день обращения, в течении часа* | Заказать | |
| МРТ грудного отдела позвоночника | в день обращения, в течении часа* | Заказать | |
| МРТ исследование одного сустава | в день обращения, в течении часа* | Заказать | |
| МРТ исследование стопы | в день обращения, в течении часа* | Заказать | |
| МРТ исследоование кисти | в день обращения, в течении часа* | Заказать | |
| МРТ копчика | в день обращения, в течении часа* | Заказать | |
| МРТ краниовертебрального перехода | в день обращения, в течении часа* | Заказать | |
| МРТ крестца и копчика | в день обращения, в течении часа* | Заказать | |
| МРТ крестцово-подвздошных сочленениий | в день обращения, в течении часа* | Заказать | |
| МРТ мягких тканей | в день обращения, в течении часа* | Заказать | |
| МРТ органов брюшной полости | в день обращения, в течении часа* | Заказать | |
| МРТ органов забрюшинного пространства (почки, надпочечники) | в день обращения, в течении часа* | Заказать | |
| МРТ органов малого таза | в день обращения, в течении часа* | Заказать | |
| МРТ органов средостения | в день обращения, в течении часа* | Заказать | |
| МРТ печени | в день обращения, в течении часа* | Заказать | |
| МРТ плюсне-фаланговых суставов | в день обращения, в течении часа* | Заказать | |
| МРТ поджелудочной железы | в день обращения, в течении часа* | Заказать | |
| МРТ пояснично-крестцового отдела позвоночника | в день обращения, в течении часа* | Заказать | |
| МРТ придаточных пазух носа | в день обращения, в течении часа* | Заказать | |
| МРТ пястно-фаланговых суставов | в день обращения, в течении часа* | Заказать | |
| МРТ тазобедренных суставов | в день обращения, в течении часа* | Заказать | |
| МРТ шейного отдела позвоночника | в день обращения, в течении часа* | Заказать | |
| МРТ щитовидной железы | в день обращения, в течении часа* | Заказать | |
| МРТ щитовидной железы и слюнных желез | в день обращения, в течении часа* | Заказать | |
| Наименование | Биоматериал | Срок | |
| Доплерография полового члена на УЗ-томографе | * | Заказать | |
| Дублексное сканирование брахиоцефальных артерий (шея+голова) | Заказать | ||
| Дублексное сканирование брахицефальных артерий (голова) | Заказать | ||
| Дублексное сканирование брахицефальных артерий (шея) | Заказать | ||
| Нейросонография (дети до 1 года) | * | Заказать | |
Обследование новорожденного до 1 мес. (нейросонография,брюшная полость,тазобедренные суставы,сердце) (нейросонография,брюшная полость,тазобедренные суставы,сердце) |
Заказать | ||
| Снимок УЗИ | Заказать | ||
| Транскраниальная доплерография | * | Заказать | |
| Транскраниальное дублексное сканирование (ТКДС) | Заказать | ||
| Транскраниальное дублексное сканирование (ТКДС) + поворотные пробы | Заказать | ||
| Триплексное сканирование почечных артерий с ЦДК | * | Заказать | |
| Триплексное сканирование почечных вен с ЦДК | * | Заказать | |
| УЗДГ беременных | Заказать | ||
| УЗДГ брюшной аорты и ее висцеральных ветвей с ЦДК | * | Заказать | |
| УЗДГ брюшной аорты и подвздошных артерий с ЦДК | * | Заказать | |
| УЗДГ нижней полой вены с ЦДК | * | Заказать | |
| УЗДГ почечных артерий с ЦДК | * | Заказать | |
| УЗДГ почечных вен с ЦДК | * | Заказать | |
| УЗДС с ЦДК артерий верхних конечностей | * | Заказать | |
| УЗДС с ЦДК артерий нижних конечностей | * | Заказать | |
| УЗДС с ЦДК брахиоцефальных артерий (БЦА) | * | Заказать | |
| УЗДС с ЦДК брюшной аорты и ее висцеральных ветвей | Заказать | ||
| УЗДС с ЦДК брюшной аорты и подвздошных артерий | Заказать | ||
| УЗДС с ЦДК вен верхних конечностей | * | Заказать | |
| УЗДС с ЦДК вен нижних конечностей | * | Заказать | |
| УЗДС с ЦДК нижней полой вены | Заказать | ||
| УЗДС с ЦДК почечных артерий | Заказать | ||
| УЗДС с ЦДК почечных вен | Заказать | ||
| УЗИ беременных 1 триместр | Заказать | ||
| УЗИ беременных 2 триместр | Заказать | ||
| УЗИ беременных 3 триместр с доплером | Заказать | ||
| УЗИ брюшной полости(печень,желчный пузырь,поджелудочная железа,селезенка, почки) | Заказать | ||
| УЗИ брюшной полости (печень, желчный пузырь, поджелудочная железа, селезенка) | * | Заказать | |
| УЗИ брюшной полости дети | Заказать | ||
| УЗИ брюшной полости с ЦДК | Заказать | ||
| УЗИ вилочковой железы | Заказать | ||
| УЗИ голеностопных суставов и стопы | * | Заказать | |
| УЗИ женских половых органов дополнительным датчиком | * | Заказать | |
| УЗИ женских половых органов ТВ датчиком с ЦДК сосудов миоматозных узлов, опухолей матки и яичников | * | Заказать | |
| УЗИ женских половых органов ТВ датчиком с ЦДК сосудов яичников, эндометрия, миометрия | * | Заказать | |
| УЗИ женских половых органов трансабдоминальным датчиком | * | Заказать | |
| УЗИ женских половых органов трансабдоминальным датчиком (врач вышей категории) | Заказать | ||
| УЗИ женских половых органов трансвагинальным датчиком | * | Заказать | |
| УЗИ женских половых органов трансвагинальным датчиком (врач высшей категории) | Заказать | ||
| УЗИ кава-фильтра с ЦДК | * | Заказать | |
| УЗИ коленных суставов | * | Заказать | |
| УЗИ коленных суставов (2 сустава) | Заказать | ||
| УЗИ лимфатических узлов дети | Заказать | ||
| УЗИ лимфоузлов | * | Заказать | |
| УЗИ локтевых суставов | * | Заказать | |
| УЗИ лучезапястных суставов и мелких суставов кистей | * | Заказать | |
| УЗИ малого таза дети | Заказать | ||
| УЗИ мелких суставов стоп | Заказать | ||
| УЗИ молочных желез | * | Заказать | |
| УЗИ молочных желез (дети) | Заказать | ||
| УЗИ мочевого пузыря с определением остаточной мочи | * | Заказать | |
| УЗИ мошонки дети | Заказать | ||
| УЗИ мошонки с доплером | * | Заказать | |
| УЗИ мягких тканей | * | Заказать | |
| УЗИ печени и желчного пузыря | * | Заказать | |
| УЗИ печени и желчного пузыря с определением функции | * | Заказать | |
| УЗИ пищевода и желудка | Заказать | ||
| УЗИ плевральной полости | Заказать | ||
| УЗИ плечевых суставов | * | Заказать | |
| УЗИ плода | * | Заказать | |
| УЗИ плода 4D с записью на DVD | * | Заказать | |
| УЗИ плода с исследованием кровотока | * | Заказать | |
| УЗИ поджелудочной железы | * | Заказать | |
| УЗИ полового члена на УЗИ-томографе | * | Заказать | |
| УЗИ почек и мочевыводящих путей с доплером дети | Заказать | ||
| УЗИ почек и надпочечников | * | Заказать | |
| УЗИ почек, надпочечников дети | Заказать | ||
| УЗИ почек, надпочечников, мочевого пузыря и предстательной железы | * | Заказать | |
| УЗИ предстательной железы и мочевого пузыря с определением остаточной мочи | * | Заказать | |
| УЗИ предстательной железы трансректальным датчиком | * | Заказать | |
| УЗИ селезенки | * | Заказать | |
| УЗИ сердца | * | Заказать | |
| УЗИ сердца дети | Заказать | ||
| УЗИ слюнной железы | Заказать | ||
| УЗИ суставов (2 суставов) дети | Заказать | ||
| УЗИ тазобедренных суставов | * | Заказать | |
| УЗИ тазобедренных суставов (дети до 1 года) | * | Заказать | |
| УЗИ тазобедренных суставов дети | Заказать | ||
| УЗИ щитовидной железы | * | Заказать | |
| УЗИ щитовидной железы дети | Заказать | ||
| УЗИ яичек | * | Заказать | |
| УЗИ яичек (старше 12 лет) | * | Заказать | |
| Фолликулометрия | * | Заказать | |
| Экстракраниальное триплексное сканирование венозной системы головного мозга | * | Заказать | |
| Наименование | Биоматериал | Срок | |
| 17-кетостероиды, моча, мг/сут | моча (суточная) | 5 рабочих дней* | Заказать |
| 17-ОН прогестерон (ИФА), кровь, нг/мл | кровь | 5 рабочих дней* | Заказать |
| D-димер, кровь, нг FEU/мл | кровь | 1 рабочий день* | Заказать |
| N-терминальный фрагмент мозгового натрийуретического пептида (NTproBNP) | кровь | 7 рабочих дней* | Заказать |
| PAPP-A (плазменный белок А, ассоциированный с беременностью) | кровь | 10 рабочих дней* | Заказать |
| АКТГ (Siemens), кровь, пг/мл | кровь | 1 рабочий день* | Заказать |
| Альбумин/креатинин, моча | 1 рабочий день* | Заказать | |
| Альдостерон (ИФА), кровь, пг/мл | кровь | 5 рабочих дней* | Заказать |
| Андростендион (Siemens), кровь, нг/мл | кровь | 10 рабочих дней* | Заказать |
| Анти-ТГ (Siemens), кровь, МЕ/мл | кровь | 1 рабочий день* | Заказать |
| Анти-ТПО (Siemens), кровь, МЕ/мл | кровь | 1 рабочий день* | Заказать |
| Антимюллеров гормон (MIS/AMH), кровь, нг/мл | кровь | 1 рабочий день* | Заказать |
| АТ к декарбоксилазе глутаминовой кислоты (AT-GAD), кровь | кровь | 14 рабочих дней* | Заказать |
| АТ к рецепторам ТТГ, кровь, МЕ/л | кровь | 10 рабочих деней* | Заказать |
| Вальпроевая кислота (Siemens), кровь, мкг/мл | кровь | 1 рабочий день* | Заказать |
| Ванилилминдальная кислота, моча, мг/24 ч | моча (суточная) | 2 рабочих дня* | Заказать |
| Витамин В12, плазма, пг/мл | кровь | 1 рабочий день* | Заказать |
| Витамин Д (25-ОН), нг/мл | * | Заказать | |
| Гликоделин, кровь, нг/мл | кровь | 14 рабочих дней* | Заказать |
| Гликозилированный гемоглобин (HbA1c), кровь, % | кровь | 5 рабочих дней* | Заказать |
| Глобулин, связывающий половые гормоны (Siemens), кровь, нМоль/л | кровь | 1 рабочий день* | Заказать |
| Гомованилиновая кислота (HVA, моча), мг/сутки | суточная моча | 1 месяц* | Заказать |
| Гомоцистеин (Siemens), кровь, мкмоль/л | Заказать | ||
| ДГЭА (дегидроэпиандростерон), кровь, ИФА, нг/мл | Заказать | ||
| Дезоксипиридинолин (ДПИД, Pyrilinks-D, Siemens), моча | моча | 1 рабочий день* | Заказать |
| Дезоксипиридинолин, нМоль/мМоль креатинина, моча | моча | 1 рабочий день* | Заказать |
| ДЭАС (дегидроэпиандостерон сульфат) (Siemens), мкг/дл | кровь | 1 рабочий день* | Заказать |
| Ингибин В, кровь, пг/мл | кровь | 1 рабочий день* | Заказать |
| Индекс свободных андрогенов | кровь | 1 рабочий день* | Заказать |
| Инсулин (Siemens), кровь, мкМЕ/мл | кровь | 1 рабочий день* | Заказать |
| Кальцитонин (Siemens), кровь, пг/мл | кровь | 1 рабочий день* | Заказать |
| Карбамазепин (Siemens), кровь, мкг/мл | кровь | 1 рабочий день* | Заказать |
| Кортизол (Siemens), кровь, мкг/дл | кровь | 1 рабочий день* | Заказать |
| Кортизол свободный, моча, мкг/сут | моча | 15 рабочих дней* | Заказать |
| Креатинин, моча, мМоль/л | моча | 1 рабочий день* | Заказать |
| КФК МВ mass (Siemens), кровь, мкг/дл | кровь | 1 рабочий день* | Заказать |
| ЛГ (Siemens), кровь, мМЕ/мл | кровь | 1 рабочий день* | Заказать |
| Лептин, кровь, нг/мл | кровь | 19 рабочих дней* | Заказать |
| Макропролактин, кровь, нг/мл | кровь | 1 рабочий день* | Заказать |
| Мета-комби (Метанефрин+Норметанефрин), моча, мг/сутки | моча (суточная) | 14 рабочих дней* | Заказать |
| Микроальбуминурия (Siemens), мкг/мин | моча | 1 рабочий день* | Заказать |
| Остеокальцин (Siemens), кровь, нг/мл | кровь | 1 рабочий день* | Заказать |
| Остеокальцин (ИФА), кровь, нг/мл | Заказать | ||
| Паратгормон (Siemens), пг/мл | кровь | 1 рабочий день* | Заказать |
| Пепсиногены I и II | кровь | 1 рабочий день* | Заказать |
| Плацентарный лактоген, кровь, мг/л | кровь | 19 рабочих дней* | Заказать |
| Прогестерон (Siemens), кровь, нг/мл | кровь | 1 рабочий день* | Заказать |
| Проинсулин, кровь, пмоль/л | кровь | 19 рабочих дней* | Заказать |
| Пролактин (Siemens), кровь, нг/мл | кровь | 1 рабочий день* | Заказать |
| Ренин + ангиотензин I | кровь | 12 рабочих дней* | Заказать |
| Ренин прямой, кровь, мкМЕ/мл | кровь | 12 рабочих дней* | Заказать |
| С-пептид (Siemens), нг/мл | кровь | 1 рабочий день* | Заказать |
| Соматомедин (инсулиноподобный фактор роста, IGF-1), сыворотка, нг/мл | кровь | 1 рабочий день* | Заказать |
| Соматотропин (Siemens), кровь, нг/мл | кровь | 1 рабочий день* | Заказать |
| Стимулирующие АТ-рТТГ , кровь, МЕ/л | кровь | 14 рабочих дней* | Заказать |
| Т3 общий (Advia Siemens), кровь, нг/дл | Кровь | 1 рабочий день* | Заказать |
| Т3 общий (Immulite Siemens), кровь, нг/дл | кровь | 1 рабочий день* | Заказать |
| Т3 свободный (Advia Siemens), кровь, пг/мл | кровь | 1 рабочий день* | Заказать |
| Т4 общий (Advia Siemens), кровь, мкг/дл | кровь | 1 рабочий день* | Заказать |
| Т4 общий (Immulite Siemens), кровь, мкг/дл | кровь | 1 рабочий день* | Заказать |
| Т4 свободный (Advia Siemens), кровь, нг/дл | кровь | 1 рабочий день* | Заказать |
| Т4 свободный (Immulite Siemens), кровь, нг/дл | кровь | 1 рабочий день* | Заказать |
| Т4 свободный (Immulite Siemens), кровь, пМоль/л | Заказать | ||
| Тестестерон свободный (Siemens), кровь, % | * | Заказать | |
| Тестостерон (Siemens), кровь, нг/дл | кровь | 1 рабочий день* | Заказать |
| Тестостерон биодоступный, кровь, % | * | Заказать | |
| Тестостерон свободный, кровь, пг/мл | кровь | 15 рабочих дней* | Заказать |
| Тиреоглобулин (Siemens), кровь, нг/мл | кровь | 1 рабочий день* | Заказать |
| Тропонин I (Siemens), кровь, нг/мл | кровь | 1 рабочий день* | Заказать |
| ТТГ (Immulite 2000 Siemens), кровь, мкМЕ/мл | кровь | 1 рабочий день* | Заказать |
ТТГ 3-я генер. (Immulite 2000 Siemens), кровь, мкМЕ/мл (Immulite 2000 Siemens), кровь, мкМЕ/мл
|
кровь | 1 рабочий день* | Заказать |
| Фолиевая кислота, плазма, нг/мл | кровь | 1 рабочий день* | Заказать |
| Фолиевая кислота, цельная кровь, нг/мл | кровь | 1 рабочий день* | Заказать |
| Фолиевая кислота, эритроциты, нг/мл | кровь | 1 рабочий день* | Заказать |
| ФСГ (Siemens), кровь, мМЕ/мл | кровь | 1 рабочий день* | Заказать |
| ХГЧ (Siemens), кровь, мМЕ/мл | кровь | 1 рабочий день* | Заказать |
| ХГЧ (Siemens), моча, мМЕ/мл | моча | 1 рабочий день* | Заказать |
| Эстрадиол (Siemens), кровь, пг/мл | кровь | 1 рабочий день* | Заказать |
Неуязвимые.
 «В Мире науки» №3, 2021
«В Мире науки» №3, 2021
Существуют ли люди, от природы не подверженные инфекционным заболеваниям? В американском блокбастере «Заражение», который многие сегодня называют пророческим, нам показывают историю распространения некоего нового природного вируса, на наших глазах поражающего огромное количество людей. Практически все умирают — вирус оказывается высококонтагиозным и патологичным. Однако один из главных героев, персонаж Мэтта Деймона, не заболел, хотя находился в самом эпицентре эпидемии. Почему он оказался не подверженным этому вирусу? Можно ли объяснить такой феномен с научной точки зрения? Действительно ли одни люди рискуют заразиться, а другим это по какой-то причине не грозит? Если да, то можно ли использовать это свойство на благо всего человечества? Эти вопросы мы задали нашим экспертам.
Виталий Васильевич Зверев, академик, заведующий кафедрой микробиологии, вирусологии и иммунологии Сеченовского университета, научный руководитель НИИ вакцин и сывороток им. И.И. Мечникова:
И.И. Мечникова:
Вы задаете непростые вопросы. Вирусов — огромное разнообразие, и мы знаем далеко не все. Но теоретически такое вполне возможно. Мы все знаем, насколько тяжелое заболевание ВИЧ- инфекция и какой серьезной проблемой становится ее широкое распространение. Но ведь известно. что есть люди, которые заражаются, но никогда не болеют. Их очень мало. Это те люди, которые имеют мутацию в гене — рецепторе к этому вирусу. Таких людей больше всего среди финно-угорских народов. У них вирус попадает в организм, но там не размножается, не заражает клетки и люди не болеют. К сожалению, это один из тех немногих случаев, которые нам известны.
Конечно, то, как люди переносят заболевание, во многом зависит от дозы заражения. Каждый вирус имеет свой уровень контагиозности. Например, для того чтобы человек заболел корью, не нужно большое количество вируса. Или та же самая ВИЧ-инфекция. Здесь очень важно, какое количество вируса попадает в организм человека, и тогда иммунная система с этим справляется или не справляется.
Наши последние исследования как раз нацелены на то, чтобы не убивать вирус, который попадает в организм человека, а ограничивать его количество, чтобы можно было создать иммунитет к нему, но при этом не болеть тяжело. Это один из возможных подходов.
А вообще здесь можно действовать на три мишени. Ведь что такое заражение вирусом? Это сам вирус; это тот рецептор, к которому вирус присоединяется; и, наконец, это иммунная система человека, который борется с этим вирусом. Для того чтобы понять, заболеет человек или нет, нужно изучить эти три позиции. Сейчас пришел новый коронавирус, а завтра придет какой-нибудь кишечный вирус или что-то еще. Каждый раз это новые пути заражения, новый источник инфекции.
Можно сколько угодно создавать противовирусные препараты, но они будут работать только против того вируса, с которым мы боремся, а с новым они уже бороться никак не смогут.
То же самое касается рецепторов. Все рецепторы вирусов имеют определенную функцию. Один из них у коронавируса, шиповидный белок-рецептор на поверхности вириона — это рецептор для связывания с ангиотензинпревращающим белком-рецептором на поверхности клеток человека. Это важнейший рецептор, который играет огромную роль в нашей жизнедеятельности. Можно, конечно, заблокировать на какое-то время эти рецепторы, но это тоже будет не очень хорошо. А можно, например, сделать упор на то, чтобы найти те точки иммунной системы, которые отвечают за развитие вируса, чтобы подавить его и одновременно вызвать небольшой иммунный ответ.
Все рецепторы вирусов имеют определенную функцию. Один из них у коронавируса, шиповидный белок-рецептор на поверхности вириона — это рецептор для связывания с ангиотензинпревращающим белком-рецептором на поверхности клеток человека. Это важнейший рецептор, который играет огромную роль в нашей жизнедеятельности. Можно, конечно, заблокировать на какое-то время эти рецепторы, но это тоже будет не очень хорошо. А можно, например, сделать упор на то, чтобы найти те точки иммунной системы, которые отвечают за развитие вируса, чтобы подавить его и одновременно вызвать небольшой иммунный ответ.
Это именно то, чем мы занимаемся вместе с Институтом общей генетики РАН и с новосибирским Институтом химической биологии и фундаментальной медицины СО РАН, с академиком В.В. Власовым. Дело в том, что ни один вирус не несет все то, что ему необходимо для развития. Он использует клеточные ферменты, белки человека, которые участвуют в регуляции иммунной системы. И можно на какое-то время прервать этот процесс, добившись таким образом того, что человек будет к этому вирусу какое-то время невосприимчив. Мы не можем надолго заблокировать компоненты иммунной системы. Но их можно будет отключить на короткий период, чтобы остановить процесс. Такие работы активно ведутся.
Мы не можем надолго заблокировать компоненты иммунной системы. Но их можно будет отключить на короткий период, чтобы остановить процесс. Такие работы активно ведутся.
Виталий Васильевич, всем нам известны истории, когда вся семья заболела, а один человек — нет. Почему такое происходит? Ему просто повезло или у него тоже какая-то мутация? Есть ли какие-то научные исследования на эту тему?
Научных исследований на эту тему было не так много, и их трудно проводить. Мы же не можем специально заразить человека и посмотреть, как он будет себя чувствовать. Но такие семейные случаи, о которых вы говорите, действительно широко известны. Это касается и коронавируса. Что мы тут знаем наверняка? Как я уже сказал, многое зависит от дозы. Далее, известно, что дети заражаются, но. как правило, не болеют или болеют очень легко. Могу сказать по своей семье — у меня двое внуков. Мы все переболели, и все по-разному, причем независимо от возраста. У кого-то были пневмонии, у кого-то — только потеря вкуса. У меня всего один день была температура. Я даже не лечился ничем, а уровень антител у меня почти такой же, как у тех, кто переболел с серьезными симптомами. А вот дети вообще бессимптомно болели, при том, что уровень иммуноглобулина G y них выше, чем у тех, кто болел тяжело. Это особенность иммунной системы, и таких людей немало.
У кого-то были пневмонии, у кого-то — только потеря вкуса. У меня всего один день была температура. Я даже не лечился ничем, а уровень антител у меня почти такой же, как у тех, кто переболел с серьезными симптомами. А вот дети вообще бессимптомно болели, при том, что уровень иммуноглобулина G y них выше, чем у тех, кто болел тяжело. Это особенность иммунной системы, и таких людей немало.
А как вы можете это объяснить? Почему так происходит? Знаю, что ваш сын болел достаточно тяжело, хотя он моложе вас.
Многое зависит от того, как человек заразился. У меня в организм, видимо, попало небольшое количество вируса, с которым моя иммунная система справилась. А у сына был очень тесный контакт с пациенткой, которая тяжело болела. У нее была коронавирусная пневмония, и он делал ей искусственное дыхание. То есть у него огромная доза вируса попала моментально, причем сразу в глубокие отделы дыхательных путей. От этого многое зависит — где вирус начинает размножаться. Плюс состояние иммунной системы. Это то, что мы сейчас и пытаемся искать: как, каким образом, какие гены включить, какие выключить, чтобы человек эту инфекцию перенес легко и даже незаметно. Это вопрос для изучения, но это очень перспективные исследования и они сейчас проводятся во всем мире.
Плюс состояние иммунной системы. Это то, что мы сейчас и пытаемся искать: как, каким образом, какие гены включить, какие выключить, чтобы человек эту инфекцию перенес легко и даже незаметно. Это вопрос для изучения, но это очень перспективные исследования и они сейчас проводятся во всем мире.
Может быть, надо изучать эти мутации, которые позволяют таким людям не болеть, и каким-то образом использовать эти знания для остальных?
Все правильно. Это надо пытаться делать. Что касается ВИЧ, мы знаем про эту мутацию. Но ее искусственно пока сделать нельзя, да и надо ли — вот вопрос. Люди с этими мутациями наверняка имеют какой-то другой дефект. Все взаимосвязано. Поэтому все научные работы в этом направлении необходимы, но очень аккуратные, чтобы, пытаясь помочь, не навредить еще больше.
Николай Иванович Брико, академик, директор Института общественного здоровья, заведующий кафедрой эпидемиологии Сеченовского университета, главный эпидемиолог Минздрава России:
Николай Иванович, сегодняшняя наша тема — устойчивость к различным возбудителям болезней. Правда ли, что у разных людей эта восприимчивость разная, а есть люди вообще невосприимчивые к тем или иным возбудителям?
Правда ли, что у разных людей эта восприимчивость разная, а есть люди вообще невосприимчивые к тем или иным возбудителям?
Да, сегодня накоплено достаточно фактического материала, который свидетельствует о гетерогенности человеческой популяции и разной восприимчивости не только к инфекционным болезням, но и вообще к любой патологии. Сегодня много данных, которые говорят о разной степени чувствительности или устойчивости к различным патогенам, с которыми сталкивается человек, как инфекционной, так и неинфекционной природы. Есть тут и гендерные различия. Мы знаем, что в этом «повинен» белок, ответственный за активацию клеточного иммунитета. Ген, обеспечивающий выработку этого белка, находится на Х-хромосоме, но, как известно, у женщин две Х-хромосомы, поэтому они могут болеть реже мужчин.
Изменчивость вирусов в значительной степени определяется адаптацией к условиям репродукции и механизмам противовирусной защиты клеток хозяина. На уровне популяций адаптация связана с особенностями типа HLA, определяющих специфичность распознавания вирусных антигенов. Генетический профиль популяции и уровень разнообразия гаплотипов HLA
не только влияют на уровень заболеваемости, но и могут определять судьбу целых этнических групп населения, их численность и показатели преждевременной смертности. В такой многонациональной стране, как Россия, этот подход должен стать основой профилактического направления практической медицины.
Генетический профиль популяции и уровень разнообразия гаплотипов HLA
не только влияют на уровень заболеваемости, но и могут определять судьбу целых этнических групп населения, их численность и показатели преждевременной смертности. В такой многонациональной стране, как Россия, этот подход должен стать основой профилактического направления практической медицины.
Есть ли какие-то данные о нынешнем коронавирусе? Кто болеет чаще?
Мы получили и продолжаем получать много информации, которая характеризует и возбудитель, и патогенез заболевания. Выстроена довольно четкая схема и лечения, и профилактики. Но многие вопросы остаются неясными, до конца не изученными, и особенно это касается вопросов генетики. Есть данные о том, что мужчины болеют тяжелее, умирают чаще. Известны возрастные различия в восприятии инфекции. Но повторяю: это та область, которую надо изучать.
Мы знаем, что есть люди, невосприимчивые к ВИЧ-инфекции. А есть ли люди, резистентные к коронавирусной инфекции?
А есть ли люди, резистентные к коронавирусной инфекции?
Мы пока не можем однозначно ответить на этот вопрос относительно COVID-19. Информации пока недостаточно. Но, скорее всего, такие люди есть. В отношении ВИЧ-инфекции есть, это известно. Этот факт связывают с определенными генами, которые кодируют рецепторные молекулы, используемые ВИЧ для проникновения в клетку. Они уже идентифицированы. Инактивация одного из генов блокирует инфицирование вирусом. Это чрезвычайно важное направление. В Китае впервые отредактирован геном человеческого эмбриона, в результате чего на свет появились генетически модифицированные близнецы. Что сделал ученый? Используя технологию CRISPR—Cas9, он удалил в эмбрионах гены, которые кодируют CCR5. Тем самым он сообщил этому эмбриону низкий риск заражения ВИЧ-инфекцией.
Но редактирование гена — это очень опасная вещь. Тот же ген CCR5 отвечает не только за восприимчивость к ВИЧ-инфекции. Оказалось, что он способствует нормальной работе лейкоцитов, и тут появляется риск заражения вирусом лихорадки Западного Нила. Вообще, использование этих технологий очень рискованно, потому что они не только удаляют этот ген, но и вносят какие-то изменения в ДНК. Это создает огромные этические проблемы, хотя, повторю, такие исследования имеют чрезвычайно большие перспективы. Устанавливая какую-то предрасположенность, мы видим, посредством каких генов это может реализоваться.
Оказалось, что он способствует нормальной работе лейкоцитов, и тут появляется риск заражения вирусом лихорадки Западного Нила. Вообще, использование этих технологий очень рискованно, потому что они не только удаляют этот ген, но и вносят какие-то изменения в ДНК. Это создает огромные этические проблемы, хотя, повторю, такие исследования имеют чрезвычайно большие перспективы. Устанавливая какую-то предрасположенность, мы видим, посредством каких генов это может реализоваться.
Как вы думаете, сколько может быть в процентном отношении людей, невосприимчивых к новой коронавирусной инфекции?
Это очень непростой вопрос, особенно применительно к новой, неизученной болезни. Однако, опираясь на исторический опыт, мы можем сказать, что даже такие мощные эпидемии, как черная оспа или легочная чума, затрагивали не всех людей. Умирало 20-30%, может быть, больше. Но остальные выживали и приобретали иммунитет. Определенная часть населения сталкивалась с возбудителем, но оставалась устойчивой к нему.
Как их вычислить? Думаю, тут надо учитывать данные, полученные в процессе реализации программ вакцинации. В человеческой популяции есть 5-10% людей, которые сильно реагируют на антигены, и есть примерно столько же слабо реагирующих. Эти данные надо учитывать, и тут мы подходим в какой-то мере к персонификации. Это один из разделов так называемой 5Р-медицины, о которой мы сейчас все чаще говорим. Предиктивная. предупредительная, предсказательная медицина, как и персонализированная, невозможна сегодня без генетических исследований.
Все люди обладают уникальной ДНК, а значит, и личным набором предрасположенностей и рисков. Предсказывать патологию и находить способы профилактики и лечения сегодня возможно только на основе генетических исследований. В последние 20-30 лет мы видим здесь фантастический взлет. Сегодня настала эпоха постгеномных исследований — это метагеномика, протеомика, транскриптомика, метаболомика, метагеномика и т.д. Известно уже более 150 лекарственных препаратов с указанием на генетический маркер. Современные достижения генетики человека делают возможным последовательное выведение системы профилактической медицины на уровень генетической персонализации.
Современные достижения генетики человека делают возможным последовательное выведение системы профилактической медицины на уровень генетической персонализации.
Как вы думаете, может ли настать момент, когда человек приходит в поликлинику к своему участковому врачу, а тот говорит: «Вам обязательно надо сделать прививку от гриппа, потому что у вас высокая предрасположенность к тяжелому течению этого заболевания; а вот от коронавирусной инфекции необязательно — у вас нет к ней склонности»?
Я верю, что наступит этот момент. И это не очень далекое будущее. Препараты, которые уже используются, влияют на гены, предрасполагающие к развитию различных онкологических заболеваний, системных поражений. Накоплено немало данных применительно к инфекционным болезням. Думаю, в ближайшем будущем у каждого из нас будет генетический паспорт, в котором будет записано, какие гены связаны с риском того или иного заболевания. Сегодня в стадии реализации находится задача генетического анализа для определения риска развития социально значимых болезней. Здесь важно определение фенотипических проявлений ранних стадий заболевания, выделение групп повышенного риска на основе индивидуальных генетических особенностей. На базе развития генетики сформировалась новая наука — эпигенетика. Ее суть заключается в возможности корректировать работу генов человека образом жизни, питанием и профилактикой. Это означает, что. приоткрыв тайны организма пациента, врач сосредоточивает усилия на профилактике вероятных заболеваний и патологий пациента взамен традиционного лечения уже существующих болезней. Это позволяет сохранять здоровье на долгие годы и отодвигать процессы старения клеток. Таким образом, медицина будущего строится на основе подбора индивидуальных процедур, программ, препаратов и доз медикаментов, которые будут эффективны для конкретного человека.
Здесь важно определение фенотипических проявлений ранних стадий заболевания, выделение групп повышенного риска на основе индивидуальных генетических особенностей. На базе развития генетики сформировалась новая наука — эпигенетика. Ее суть заключается в возможности корректировать работу генов человека образом жизни, питанием и профилактикой. Это означает, что. приоткрыв тайны организма пациента, врач сосредоточивает усилия на профилактике вероятных заболеваний и патологий пациента взамен традиционного лечения уже существующих болезней. Это позволяет сохранять здоровье на долгие годы и отодвигать процессы старения клеток. Таким образом, медицина будущего строится на основе подбора индивидуальных процедур, программ, препаратов и доз медикаментов, которые будут эффективны для конкретного человека.
Константин Валерьевич Крутовский, ведущий научный сотрудник Института общей генетики им. Н.И. Вавилова РАН. профессор кафедры геномики и биоинформатики и руководитель Научно-образовательного центра геномных исследований Сибирского федерального университета, профессор Геттингенского университета:
Константин Валерьевич, известно, что есть люди, устойчивые к тем или иным типам возбудителей инфекции. Во многом именно генетика может ответить на вопрос, каким людям грозит то или иное инфекционное заболевание, а каким нет. Как вы думаете, есть ли будущее у генетики в предсказании опасности вирусных заболеваний у конкретных людей?
Во многом именно генетика может ответить на вопрос, каким людям грозит то или иное инфекционное заболевание, а каким нет. Как вы думаете, есть ли будущее у генетики в предсказании опасности вирусных заболеваний у конкретных людей?
Это вопрос персонифицированной медицины, когда на основании индивидуальных геномов человека мы можем предсказывать предрасположенность к тем или иным болезням. Чисто гипотетически можно на основании индивидуального генома человека сделать вывод, есть ли склонность к каким-либо аутоиммунным заболеваниям или пониженный иммунитет к разным патогенным. Такие исследования уже активно проводятся, и, конечно, в перспективе на их основе можно будет делать надежные предсказания.
Но тут есть еще один важный вопрос — врожденная устойчивость к болезням. Почему одни люди не заболевают даже при контакте с вирусами или болеют в очень легкой форме, а другие нет? В чем причина такого феномена? Может быть, человек когда-то уже контактировал с этим вирусом? Существует врожденный клеточный иммунитет.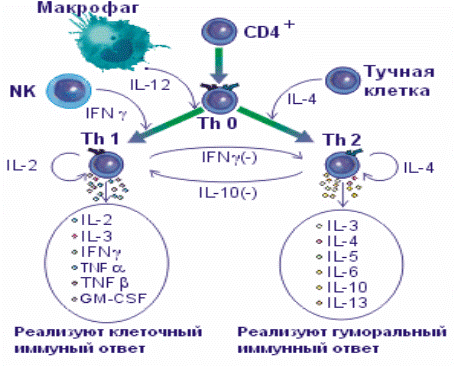
Есть очень интересные исследования по эпигеномике, говорящие о том, что приобретенная устойчивость тоже может передаваться — необязательно на уровне замен нуклеотидов в ДНК, но через эпигенетические механизмы, через обратимые химические модификации нуклеотидов и хроматина.
С чем это связано? Врожденное ли это? Может быть, это обусловлено также тем, что у человека уже известны четыре вида коронавирусов — два альфа- и два бета-коронавируса, с которыми он давно и относительно мирно сосуществует? Причем бета-коронавирусы — это группа, к которой принадлежит SARS CoV-2. Они уже есть у человека, давно открыты и обычно вызывают легкие респираторные заболевания. То есть они адаптировались к организму человека. И, возможно, тот иммунитет, который к ним вырабатывается, особенно у тех людей, которые переболели этими коронавирусами, включает как некий бонус устойчивость к новому для человека коронавирусу.
Это одно из возможных научных объяснений, но этот вопрос надо изучать.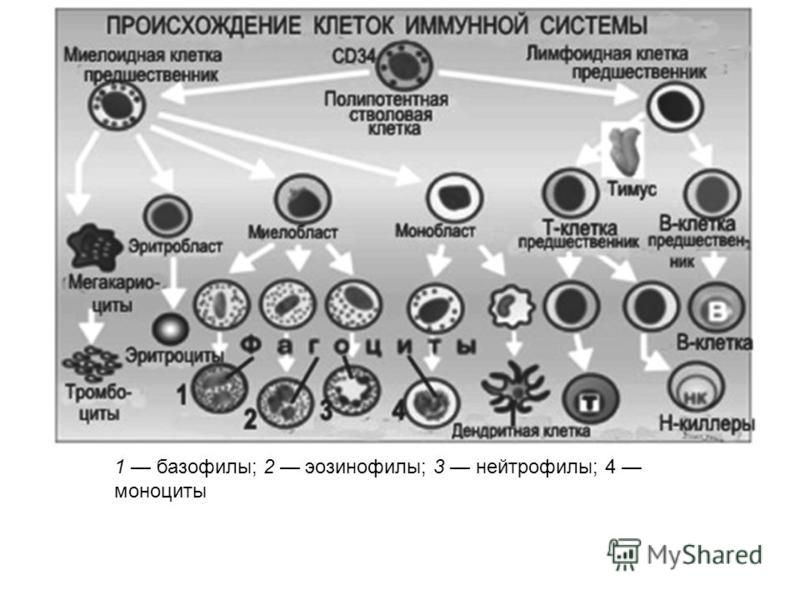 Если предположение верно, у нас появляется надежда, что и нынешний вирус тоже адаптируется. Вирусу вообще невыгодно убивать хозяина, потому что с его гибелью он погибает сам. Эволюционно он заинтересован наносить минимальный вред, но при этом быть максимально инфекционным, то есть заразным. Поэтому естественный отбор отбирает те мутации, которые снижают его летальность, патогенность, но увеличивают его контагиозность. И у нас есть много примеров таких вирусов: некоторые аденовирусы, коронавирусы, вирусы гриппа и др.
Если предположение верно, у нас появляется надежда, что и нынешний вирус тоже адаптируется. Вирусу вообще невыгодно убивать хозяина, потому что с его гибелью он погибает сам. Эволюционно он заинтересован наносить минимальный вред, но при этом быть максимально инфекционным, то есть заразным. Поэтому естественный отбор отбирает те мутации, которые снижают его летальность, патогенность, но увеличивают его контагиозность. И у нас есть много примеров таких вирусов: некоторые аденовирусы, коронавирусы, вирусы гриппа и др.
Но ведь не со всеми вирусами так произошло.
Да, не со всеми. С вирусами оспы, кори, полиомиелита этого не случилось. Поэтому надо вакцинироваться. Я считаю, что вакцинирование — это гражданский долг каждого, потому что защищаешь не только себя, но и других. Это помогает создать так называемый популяционный иммунитет, который позволит прервать текущую пандемию. Чем быстрее и чем больше людей переболеет или вакцинируется, тем лучше. Но лучше не болеть, поскольку это вызывает и тяжелые осложнения, и жертвы.
Но лучше не болеть, поскольку это вызывает и тяжелые осложнения, и жертвы.
Знаю, что сейчас очень много колеблющихся. Люди боятся вакцинации, не доверяют ей. Думаю, всем, кто вакцинируется, необходимо давать специальную медицинскую страховку на случай возможных осложнений. Они редкие, но могут быть. Такая страховка важна, чтобы была медицинская гарантированная помощь, какое-то финансирование или даже существенная компенсация. Подобную программу должно предусмотреть государство, заинтересованное в массовой вакцинации.
Верно ли я понимаю, что развитие вирусологии без генетики сегодня невозможно?
Да, все верно. Без генетики сегодня ни одна биологическая дисциплина не может нормально развиваться, поскольку это основа основ: ведь гены регулируют все функции организма. Генетика, я считаю, — одна из ключевых биологических дисциплин. Это наука о наследственности и изменчивости у всех живых организмов, где есть гены, есть ДНК, есть генетический контроль, где совершаются мутации, возникает изменчивость, а на ее основе происходят адаптация и эволюция. Понимание этих механизмов, адаптации и эволюции, того, как вирус эволюционирует, без генетики, конечно, невозможно. Как и создание вакцин, имеющих минимальное количество побочных последствий, невозможно себе представить без генетики.
Понимание этих механизмов, адаптации и эволюции, того, как вирус эволюционирует, без генетики, конечно, невозможно. Как и создание вакцин, имеющих минимальное количество побочных последствий, невозможно себе представить без генетики.
Для создания таких современных, высокоэффективных и безопасных вакцин надо знать устройство генома, его последовательности, а сейчас это можно делать на уровне генетического текста, прочитывая его с помощью секвенирования. Сегодня научились выделять именно те фрагменты белка, которые вызывают наиболее сильный иммунный ответ, но минимальные побочные эффекты. Это так называемые пептидные вакцины, например «ЭпиВакКорона», разработанная в центре «Вектор». Их не надо путать с так называемыми векторными вакцинами, которые используют другой принцип. Благодаря молекулярной генетике, геномике и генной инженерии в этой области также достигнут громадный прогресс.
Одна из таких векторных вакцин — наш «Спутник V». В основе этой вакцины — модифицированный и неспособный к репликации относительно безопасный для человека аденовирус, в него встроен S-ген от SARS—CoV-2,
кодирующий шиповидный белок, на который образуются антитела и формируется иммунитет.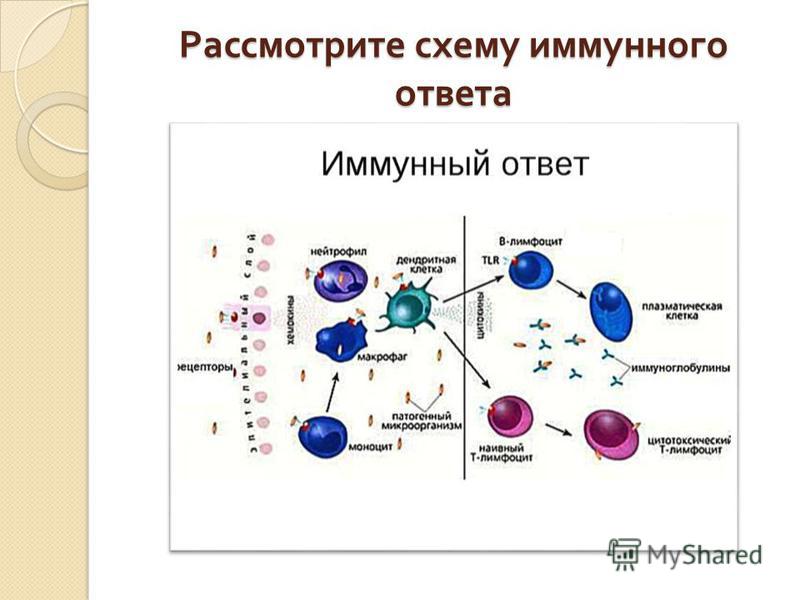 В данном случае аденовирус служит только носителем, вектором, призванным доставить ген коронавируса внутрь клетки, где он будет экспрессироваться, и на него будут нарабатываться антитела, таким образом формируя иммунитет.
В данном случае аденовирус служит только носителем, вектором, призванным доставить ген коронавируса внутрь клетки, где он будет экспрессироваться, и на него будут нарабатываться антитела, таким образом формируя иммунитет.
Есть еще более продвинутые вакцины на основе только одного или двух генов SARS—CoV-2 в виде матрицы мРНК, когда эта матрица внутри липидной наночастицы через инъекцию вводится в организм и затем проникает внутрь клетки, где идет ее считывание и происходит синтез специфичного белка, характерного для коронавируса. На него в результате и формируется иммунитет.
Думаю, в будущем наука и медицина придут к тому, что человек сможет узнать, нужна ли ему вакцинация от той или иной вирусной инфекции (и какую вакцину ему нужно предпочесть) или ему эта болезнь не грозит, потому что у него есть врожденный или приобретенный иммунитет к ней. А пока наука не научилась предсказывать это быстро и точно, необходимо массово вакцинироваться.
Беседовала Наталия Лескова
Фото: scientificrussia.ru
Ссылка на публикацию: scientificrussia.ru
Генетический контроль иммунного ответа и восприимчивости к инфекционным заболеваниям
Известно, что как генетические, так и негенетические переменные влияют по величине и широте на иммунный ответ на возбудителей инфекционных заболеваний. Открытие и проверка генетических детерминант хозяев и патогенов имеют решающее значение для лучшего понимания основ восприимчивости к инфекционным заболеваниям и борьбы с ними. Взаимодействие этих объединенных, но противоположных множественных, различных факторов приводит к впечатляющему динамическому фенотипическому разнообразию у хозяев.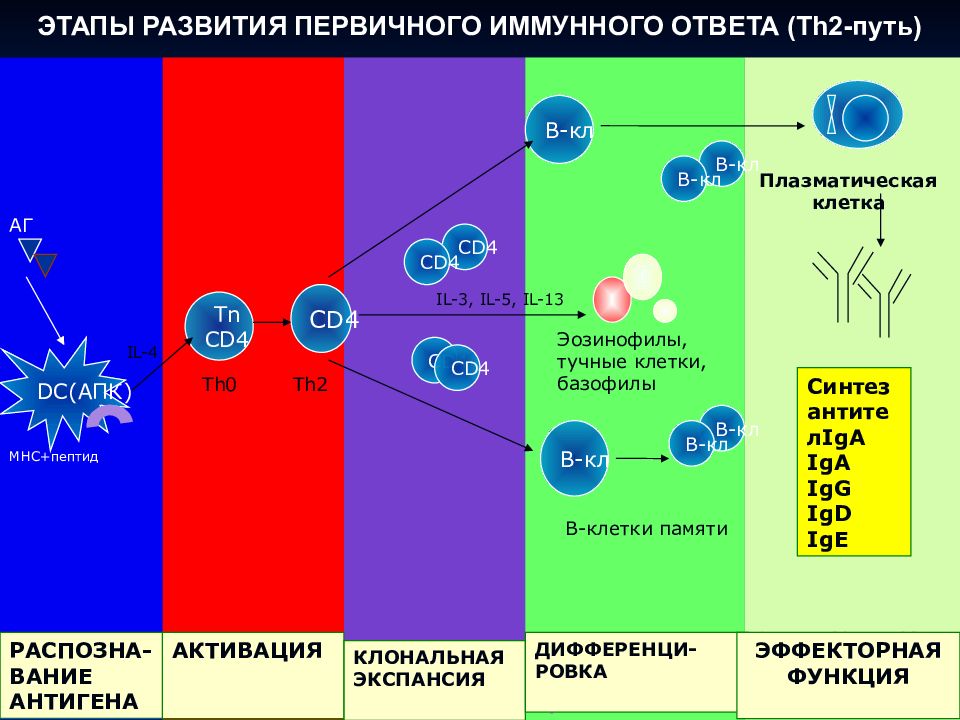 Точные механизмы, лежащие в основе наблюдаемых индивидуальных различий в контроле, устойчивости и / или восприимчивости к инфекционным заболеваниям, полностью не изучены.
Точные механизмы, лежащие в основе наблюдаемых индивидуальных различий в контроле, устойчивости и / или восприимчивости к инфекционным заболеваниям, полностью не изучены.
В этом специальном выпуске собраны три оригинальные исследовательские работы и пять обзоров, направленных на стимулирование будущих исследовательских усилий по медицинскому и ветеринарному применению генетических вариаций с четко определенной генетической ролью в иммунном ответе хозяина и восприимчивости к инфекционным заболеваниям. В целом, в этой области исследований преобладала популяционная генетика генов-кандидатов посредством небольших исследований ассоциаций индивидуальной восприимчивости.Наблюдаемые межиндивидуальные различия частично обусловлены модифицирующими риск полиморфизмами (редкими и распространенными) генов-агонистов и антагонистов врожденного и адаптивного иммунных ответов. Существуют убедительные доказательства связи с контролем или снижением восприимчивости к инфекционным агентам или прогрессирования заболевания для некоторых вариантов гена хозяина.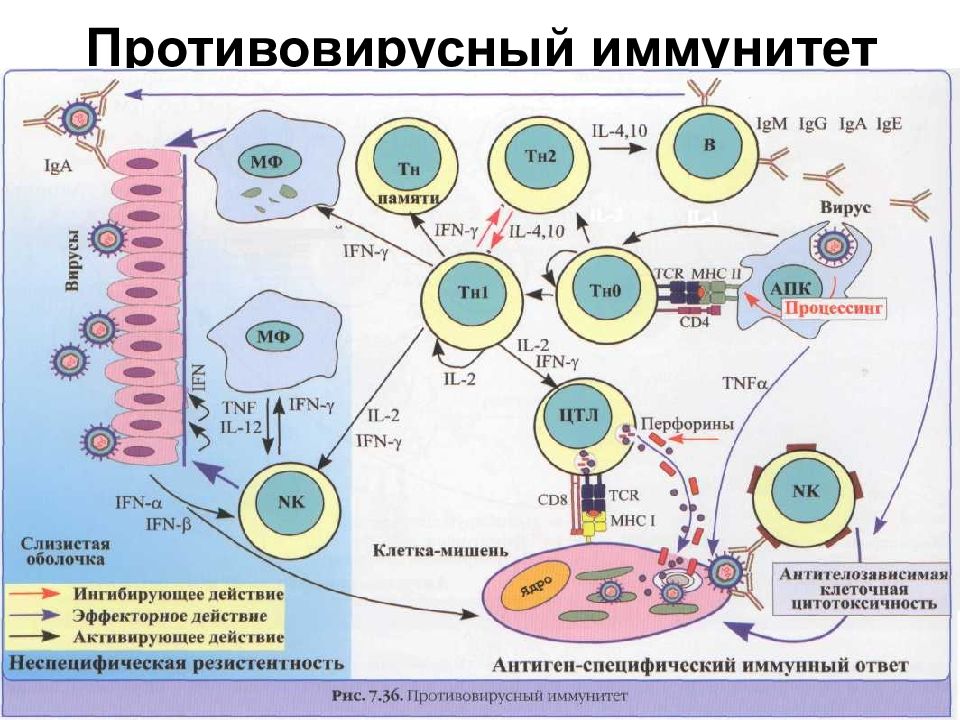 Перспектива состоит в том, что в этой области будут полезны как мелкомасштабные исследования реплик, так и исследования ассоциаций в масштабе всего генома (GWAS), во-первых, для проверки статистической силы наблюдаемых ассоциаций в различных популяционных фонах, а во-вторых, для обнаружения значимых ассоциаций с недавно исследованными вариантами с трансляционным потенциалом на практике. тесты.
Перспектива состоит в том, что в этой области будут полезны как мелкомасштабные исследования реплик, так и исследования ассоциаций в масштабе всего генома (GWAS), во-первых, для проверки статистической силы наблюдаемых ассоциаций в различных популяционных фонах, а во-вторых, для обнаружения значимых ассоциаций с недавно исследованными вариантами с трансляционным потенциалом на практике. тесты.
А. В. Марангон и др. изучили ассоциацию генов HLA класса I, HLA класса II и KIR , а также однонуклеотидные полиморфизмы (SNP) в генах цитокинов ( IFNG , IL6 , IL10 , TGFB1 , и TN ) с риском заболевания шейки матки, связанного с вирусом папилломы человека (ВПЧ), в двух группах субъектов: 79 неродственных смешанных бразильских женщин с положительным результатом теста на канцерогенный ВПЧ высокого риска с цервикальной интраэпителиальной неоплазией 3 степени (CIN3) и 150 ВПЧ. -отрицательные женщины одной национальности.Наиболее важные результаты этого исследования: (i) ни один из четырнадцати генов KIR , псевдогена KIR , аллелей HLA класса I / II (или их комбинации гаплотипов) не был существенно связан с заболеванием шейки матки, связанным с ВПЧ; (ii) гаплотип HLA-A * 02-B * 51 был связан с устойчивостью к заболеванию шейки матки, связанному с ВПЧ; (iii) генотип IFNG + 874A / A был связан с восприимчивостью к инфекции ВПЧ.
Z.-Q. Feng et al. проверили, вовлечены ли индуцированные ретиноевой кислотой ген I (RIGI) -подобные рецепторы (RLR) — обнаружение вирусов и пути противовирусного ответа против РНК-вирусов в резистентность к вирусу болезни Марека (MDV), ДНК-вирусу, у кур, и есть ли резистентность На заражение влияет генетический фон кур.Авторы экспериментально заразили две породы кур (бройлеры экономической линии AA и местные горные куры Эрланга) для сравнительного мониторинга паттернов экспрессии РНК гена 5, связанного с дифференцировкой меланомы ( MDA-5, ), фактора транскрипции 3, регулирующего интерферон ( IRF- 3 ), IFN- α , и IFN- β с помощью ПЦР в реальном времени на общих кДНК из различных тканей. Наиболее важные результаты этого исследования: (i) горная порода кур Эрланг, но не порода экономичной линии AA, выжила после заражения MDV при небольшом усилении контроля вирусной нагрузки; (ii) Экспрессия РНК для MDA-5 , IRF-3 , IFN- α и IFN- β увеличивалась до разных величин после заражения у обеих пород, но наблюдались противоположные величины у разные ткани.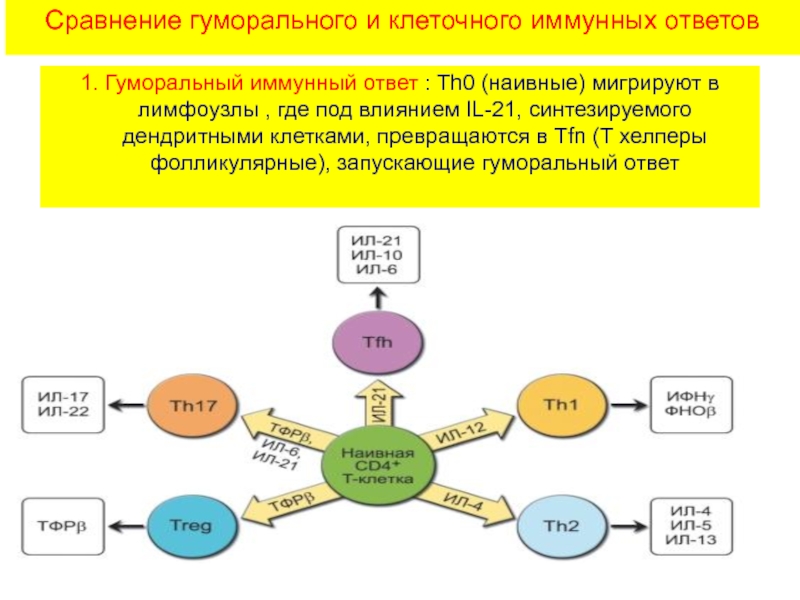 Перспектива авторов состоит в том, что RLR-опосредованный противовирусный путь участвует в обнаружении и ответе против MDV.
Перспектива авторов состоит в том, что RLR-опосредованный противовирусный путь участвует в обнаружении и ответе против MDV.
A.C. Leandro et al. опросили IFNG + 874 * T / A и NOS2A -954 * G / C SNP в исследовании случай-контроль (/), чтобы оценить их роль в предрасположенности к туберкулезу и определить, влияют ли эти варианты на уровни нитритов и нитритов. радикал оксида азота в сыворотке. Наиболее значительный результат этого исследования заключается в том, что ни вариант аллеля, ни их комбинация не были значимо связаны ни с риском развития туберкулеза, ни с различиями в уровнях секреции радикалов оксида азота в смешанной исследуемой подгруппе населения Бразилии.
C. M. Ayo et al. предоставил исчерпывающий обзор ассоциаций между полиморфизмами в HLA класса I / II, KIR , цитокинов ( CCL2 , CXCL9 , IFNG , IL1B , IL12B , IL4
, IL4 , IL4 , IL4 , IL4 IL10 , LTA , TGFB1 и TNF ), гены цитокинового рецептора ( CCR5 , IL4R ) и фактор сплайсинга BAT1 (DDX39B) ген и возникновение, тяжесть и клинические формы болезнь (американский трипаносомоз).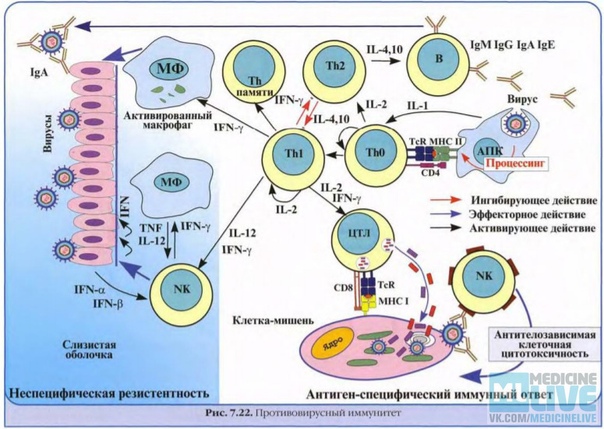 Похвальным вкладом этого обзора является отчет о статистической значимости (значениях) ассоциаций из различных исследований, наблюдаемых между аллелями и гаплотипами HLA и различными клиническими формами заболевания (хроническая кардиомиопатия Шагаса, пищеварительная форма, а также смешанные и их комбинации) . Несмотря на многочисленные исследования сцепления и ассоциации, проведенные для обнаружения генетических вариантов хозяина, участвующих в иммунопатогенезе болезни Шагаса, причинная связь восприимчивости остается проблемой.
Похвальным вкладом этого обзора является отчет о статистической значимости (значениях) ассоциаций из различных исследований, наблюдаемых между аллелями и гаплотипами HLA и различными клиническими формами заболевания (хроническая кардиомиопатия Шагаса, пищеварительная форма, а также смешанные и их комбинации) . Несмотря на многочисленные исследования сцепления и ассоциации, проведенные для обнаружения генетических вариантов хозяина, участвующих в иммунопатогенезе болезни Шагаса, причинная связь восприимчивости остается проблемой. L. R. Jarduli et al. внес полезный обзор о роли генов HLA, KIR, и MICA , а также об общих полиморфизмах про- и противовоспалительных цитокинов ( IFNG , IL1B , IL12B , IL4 , IL6 , IL10 , TNF , и LTA ) и рецептор цитокинов ( IFNGR1 , IL12RB1 и IL12RB2 ), BAT1 и BTNLyro ассоциированный ) ген устойчивости или восприимчивости к инфекции Mycobacterium leprae , а также клиническое течение и различные проявления лепры. Достойным похвалы аспектом этого обзора является охват по клиническим формам болезни (мультибациллярная, лепроматозная, пограничная, туберкулоидная и узловатая эритема лепрозная) с включением статистической значимости (значений) ассоциаций, наблюдаемых в исследованиях близнецов, анализах сегрегации и т. Д. семейные исследования сцепления и ассоциации, исследования ассоциации генов-кандидатов и, совсем недавно, GWAS. Интересно, что некоторые комбинации генов HLA и KIR оказывали благоприятные или неблагоприятные взаимодействия с противоположными (активацией или ингибированием) эффектами на NK-клетки и инфицированные клетки-хозяева, что подчеркивает взаимодействие генетических факторов хозяина в патогенезе.
Достойным похвалы аспектом этого обзора является охват по клиническим формам болезни (мультибациллярная, лепроматозная, пограничная, туберкулоидная и узловатая эритема лепрозная) с включением статистической значимости (значений) ассоциаций, наблюдаемых в исследованиях близнецов, анализах сегрегации и т. Д. семейные исследования сцепления и ассоциации, исследования ассоциации генов-кандидатов и, совсем недавно, GWAS. Интересно, что некоторые комбинации генов HLA и KIR оказывали благоприятные или неблагоприятные взаимодействия с противоположными (активацией или ингибированием) эффектами на NK-клетки и инфицированные клетки-хозяева, что подчеркивает взаимодействие генетических факторов хозяина в патогенезе.
F. Celsi et al. предоставил просвещенный отчет о планах регуляции экспрессии генов, разработанных вирусом иммунодефицита человека типа 1 (ВИЧ-1), чтобы избежать иммунного ответа против инфицированных клеток, с уделением особого внимания взаимодействию классического класса I HLA-C и неклассического класса I HLA-G аллели и генотипы, связанные с эффективным контролем репликации вируса и с медленным прогрессированием до СПИДа. Осознание того, что генетические фоновые различия (т. Е. Этническая принадлежность) в распределении аллелей HLA-G (т. Е. HLA-G * 0105N) могут влиять на результаты (защита / восприимчивость) ассоциативных исследований, настоятельно требует исследований реплик.Авторы далее оценили недавно обнаруженные малые некодирующие РНК (miRNAs), зашифрованные в геноме хозяина, для подавления экспрессии генов HLA-C и HLA-G посредством РНК-интерференции в 3′-нетранслируемых областях (UTR). Обсуждались возможные эффекты SNP на HLA-C и HLA-G 3 ‘UTR на связывание miRNA, а также перспективы использования сигнатур экспрессии miRNA в качестве биомаркера прогрессирования заболевания. По мнению авторов, оценка воздействия таких регуляторных механизмов на инфицирование ВИЧ-1, его репликацию и прогрессирование до СПИДа позволит выявить новые терапевтические цели.Перспективы существования противодействующей miRNA, зашифрованной (предположительно) в геноме ВИЧ-1, открывают путь к беспрецедентным подходам.
Осознание того, что генетические фоновые различия (т. Е. Этническая принадлежность) в распределении аллелей HLA-G (т. Е. HLA-G * 0105N) могут влиять на результаты (защита / восприимчивость) ассоциативных исследований, настоятельно требует исследований реплик.Авторы далее оценили недавно обнаруженные малые некодирующие РНК (miRNAs), зашифрованные в геноме хозяина, для подавления экспрессии генов HLA-C и HLA-G посредством РНК-интерференции в 3′-нетранслируемых областях (UTR). Обсуждались возможные эффекты SNP на HLA-C и HLA-G 3 ‘UTR на связывание miRNA, а также перспективы использования сигнатур экспрессии miRNA в качестве биомаркера прогрессирования заболевания. По мнению авторов, оценка воздействия таких регуляторных механизмов на инфицирование ВИЧ-1, его репликацию и прогрессирование до СПИДа позволит выявить новые терапевтические цели.Перспективы существования противодействующей miRNA, зашифрованной (предположительно) в геноме ВИЧ-1, открывают путь к беспрецедентным подходам.
VC Vieira и MA Soares представили новую оценку зависимой или независимой роли редактирования последовательности ДНК и / или РНК семейства цитидин дезаминаз APOBEC человека в подавлении врожденного иммунитета вирусных инфекций (включая EBV, HBV, HCV, HIV-1).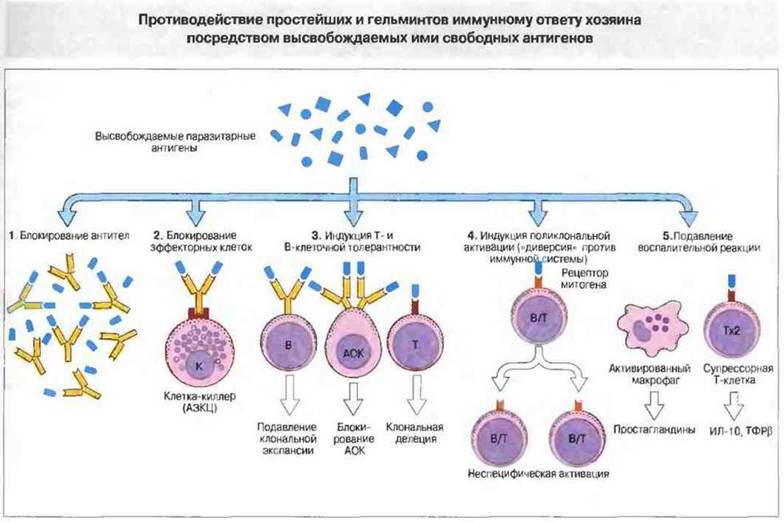 , HPV, HSV-1 и HTLV), а также противодействующие вирусные стратегии. Похвальным аспектом этого обзора является всестороннее и проницательное сравнение исследований полиморфизмов в генах APOBEC3 и их ассоциаций с восприимчивостью к вирусным инфекциям.Перспектива авторов заключается в том, что усиление активности APOBEC3 представляет собой новый подход к разработке противовирусных препаратов, несмотря на осознание того, что APOBEC3-опосредованная гипермутирующая вирусная ДНК / РНК может увеличить генетическую диверсификацию вируса и, таким образом, появление ускользнувших от вирусов и / или устойчивых к лекарствам вариантов. .
, HPV, HSV-1 и HTLV), а также противодействующие вирусные стратегии. Похвальным аспектом этого обзора является всестороннее и проницательное сравнение исследований полиморфизмов в генах APOBEC3 и их ассоциаций с восприимчивостью к вирусным инфекциям.Перспектива авторов заключается в том, что усиление активности APOBEC3 представляет собой новый подход к разработке противовирусных препаратов, несмотря на осознание того, что APOBEC3-опосредованная гипермутирующая вирусная ДНК / РНК может увеличить генетическую диверсификацию вируса и, таким образом, появление ускользнувших от вирусов и / или устойчивых к лекарствам вариантов. .
J. Malogajski et al. обобщил данные о высоком трансляционном потенциале основных геномных и генетических исследований хозяина для трех агентов, передающихся половым путем (ВИЧ-1, Chlamydia trachomatis и HPV), в приложениях общественного здравоохранения и помощи, включая диагностику, лечение и профилактику заболеваний.Авторы утверждали, что существует значительный разрыв между открытием функциональных генетических полиморфизмов и эффективным внедрением практических приложений, что указывает на плодородную почву для исследований в области фармакогеномики и персонализированной медицины. Фактически, готового к продаже приложения, включающего генетические / геномные тесты на заболевания, вызываемые этими инфекционными агентами, в настоящее время нет. Перспективы внедрения рутинных скрининговых тестов на эпигенетические (метилирование ДНК) маркеры, которые, как известно, связаны с интраэпителиальной неоплазией шейки матки высокой степени и раком шейки матки, соответствуют ожиданиям значительного сокращения количества ненужных обращений к гинекологам.Аналогичным образом ожидается, что введение TLR4 SNP-типирования и TLR2 гаплотипов для идентификации вариантов или гаплотипов, связанных либо с повышенным риском патологии маточных труб, либо с защитой от патологии маточных труб, соответственно, обеспечит более точный диагноз Chlamydia trachomatis вызывая субфертильность.
Фактически, готового к продаже приложения, включающего генетические / геномные тесты на заболевания, вызываемые этими инфекционными агентами, в настоящее время нет. Перспективы внедрения рутинных скрининговых тестов на эпигенетические (метилирование ДНК) маркеры, которые, как известно, связаны с интраэпителиальной неоплазией шейки матки высокой степени и раком шейки матки, соответствуют ожиданиям значительного сокращения количества ненужных обращений к гинекологам.Аналогичным образом ожидается, что введение TLR4 SNP-типирования и TLR2 гаплотипов для идентификации вариантов или гаплотипов, связанных либо с повышенным риском патологии маточных труб, либо с защитой от патологии маточных труб, соответственно, обеспечит более точный диагноз Chlamydia trachomatis вызывая субфертильность.
Мы ожидаем, что статьи в этом специальном выпуске повысят восприимчивость к открытию и применению генетических вариантов в биотехнологии, диагностике и лечении инфекционных заболеваний как человека, так и животных.
Энрике Медина-Акоста
Хелдер Такаши Имото Накая
Алессандра Понтилло
Регина Селия де Соуза Кампос Фернандес
Генетический контроль врожденного иммунного ответа | BMC Immunology
Джейнвей К.А., Меджитов Р: Введение: Роль врожденного иммунитета в адаптивном иммунном ответе. Семинары по иммунологии. 1998, 10: 349-350. 10.1006 / smim.1998.0142.
Артикул PubMed Google Scholar
Адерем А., Улевич Р.Дж.: Толл-подобные рецепторы в индукции врожденного иммунного ответа. Природа. 2000, 406: 782-787. 10.1038 / 35021228.
CAS Статья PubMed Google Scholar
Опал С.М., Глюк Т: Эндотоксин как мишень для лекарства. Реанимационная медицина. 2003, 31: S57-S64. 10.1097 / 00003246-200301001-00009.
CAS Статья PubMed Google Scholar
Arbor NC, Lorenz E, Schutte BC, Zabner J, Kline JN, Jones M, Frees KL, Watt JL, Schwartz DA: мутации TLR4 связаны с гипореактивностью эндотоксина у людей. Природа Генетика. 2000, 25: 187-191. 10.1038 / 76048.
CAS Статья PubMed Google Scholar
Шварц Д.А.: Генетика врожденного иммунитета. Грудь. 2002, 121: 62С-68С. 10.1378 / Chess.121.3_suppl.62S.
CAS Статья PubMed Google Scholar
Kiechl S, Lorenz E, Reindl M, Wiedermann CJ, Oberhollenzer F, Bonora E, Willeit J, Schwartz DA: Толл-подобные полиморфизмы рецептора 4 и атерогенез. Медицинский журнал Новой Англии. 2002, 347: 185-192. 10.1056 / NEJMoa012673.
CAS Статья PubMed Google Scholar
Болдрик Дж. С., Ализаде А. А., Дин М., Дудой С., Лю С. Л., Белчер С. Е., Ботштейн Д., Штаудт Л. М., Браун П. О., Релман Д. А. Стереотипные и специфические программы экспрессии генов в ответах на бактерии врожденного иммунитета человека.PNAS. 2002, 99: 972-977. 10.1073 / pnas.231625398.
А. Стереотипные и специфические программы экспрессии генов в ответах на бактерии врожденного иммунитета человека.PNAS. 2002, 99: 972-977. 10.1073 / pnas.231625398.
PubMed Central CAS Статья PubMed Google Scholar
Nau GJ, Richmond JFL, Schlesinger A, Jennings EG, Lander ES, Young RA: Программы активации человеческих макрофагов, индуцированные бактериальными патогенами. PNAS. 2002, 99: 1503-1508. 10.1073 / pnas.022649799.
PubMed Central CAS Статья PubMed Google Scholar
Rosenberger CM, Scott MG, Gold MR, Hancock REW, Finlay BB: Инфекция Salmonella typhimurium и стимуляция липополисахаридами вызывают аналогичные изменения в экспрессии генов макрофагов. J Immunol. 2000, 164: 5894-5904.
CAS Статья PubMed Google Scholar
Gao JJ, Diesl V, Wittmann T, Morrison DC, Ryan JL, Vogel SN, Follettie MT: Регулирование экспрессии генов в макрофагах мыши, стимулированных бактериальной cpG-ДНК и липополисахаридом. Журнал биологии лейкоцитов. 2002, 72: 1234-1245.
Журнал биологии лейкоцитов. 2002, 72: 1234-1245.
CAS PubMed Google Scholar
Раваси Т., Уэллс Калифорния, Форест А., Андерхилл Д.М., Уэйнрайт Б.Дж., Адерем А., Гриммонд С., Хьюм Д.А.: Генерация разнообразия в врожденной иммунной системе: неоднородность макрофагов возникает из-за генетической автономной транскрипционной вероятности индивида. Индуцибельные гены. Журнал иммунологии. 2002, 168: 44-50.
CAS Статья PubMed Google Scholar
Govoni G, Vidal S, Gauthier S, Skamene E, Malo D, Gros P: Локус Bcg / Ity / Lsh: генетический перенос устойчивости к инфекциям у мышей C57BL / 6J, трансгенных по аллелю Nramp1 Gly169. Заражение иммунной. 1996, 64: 2923-2929.
PubMed Central CAS PubMed Google Scholar
Wardrop SL, Wells C, Ravasi T, Hume DA, Richardson DR: Индукция Nramp2 в активированных макрофагах мыши не связана с регуляцией Nramp1, классических воспалительных генов и генов, участвующих в метаболизме железа. J Leukoc Biol. 2002, 71: 99-106.
J Leukoc Biol. 2002, 71: 99-106.
CAS PubMed Google Scholar
Полторак А, Хе Х, Смирнова I, Лю М.Ю., Хаффель К.В., Ду Х, Бердвелл Д., Алехос Э., Сильва М., Галанос С., Фройденберг М., Риккарди-Кастаньоли П., Лейтон Б., Бейтлер Б. Дефектная передача сигналов LPS у мышей C3H / HeJ и C57BL / 10ScCr: мутации в гене Tlr4. Наука. 1998, 282: 2085-2088. 10.1126 / science.282.5396.2085.
CAS Статья PubMed Google Scholar
Xu Y, Tao X, Shen B, Horng T, Medzhitov R, Manley JL, Tong L: Структурная основа передачи сигнала доменами рецепторов Toll / интерлейкина-1. Природа. 2000, 408: 111-115. 10.1038 / 35047056.
CAS Статья PubMed Google Scholar
Hirschfeld M, Ma Y, Weis JH, Vogel SN, Weis JJ: Передний край: повторная очистка липополисахарида устраняет передачу сигналов через толл-подобные рецепторы человека и мыши 2. J Immunol. 2000, 165: 618-622.
J Immunol. 2000, 165: 618-622.
CAS Статья PubMed Google Scholar
Wells CA, Ravasi T, Sultana R, Yagi K, Carinci P, Bono H, Faulkner G, Okazaki Y, Quackenbush J, Hume DA, Members RGG, Lyons PA: продолжающееся открытие транскрипционных единиц, экспрессируемых в клетках линии мононуклеарных фагоцитов мыши. Геномные исследования.
Sweet MJ, Campbell CC, Sester DP, Xu D, McDonald RC, Stacey KJ, Hume DA, Liew FY: Колониестимулирующий фактор-1 подавляет ответы на ДНК CpG и экспрессию Toll-подобного рецептора 9, но Усиливает реакцию на липополисахарид в макрофагах мышей.Журнал иммунологии. 2002, 168: 392-399.
CAS Статья PubMed Google Scholar
Miki R, Kadota K, Bono H, Mizuno Y, Tomaru Y, Carninci P, Itoh M, Shibata K, Kawai J, Konno H, Watanabe S, Sato K, Tokusumi Y, Kikuchi N, Ishii Y , Hamaguchi Y, Nishizuka I, Goto H, Nitanda H, Satomi S, Yoshiki A, Kusakabe M, DeRisi JL, Eisen MB, Iyer VR, Brown PO, Muramatsu M, Shimada H, Okazaki Y, Hayashizaki Y: определение развития и метаболизма пути in vivo путем профилирования экспрессии с использованием набора RIKEN из 18 816 полноразмерных обогащенных массивов кДНК мыши. Труды Национальной академии наук США. 2001, 98: 2199-2204. 10.1073 / pnas.041605498.
Труды Национальной академии наук США. 2001, 98: 2199-2204. 10.1073 / pnas.041605498.
CAS Статья Google Scholar
Тиблемонт Н., Райт С.Д.: Транспорт бактериального липополисахарида в аппарат Гольджи. Журнал экспериментальной медицины. 1999, 190: 523-534. 10.1084 / jem.190.4.523.
PubMed Central CAS Статья PubMed Google Scholar
Shurety W, Merino-Trigo A, Brown D, Hume DA, Stow JL: Локализация и перенос фактора некроза опухоли альфа в макрофагах после Гольджи. J Interferon Cytokine Res. 2000, 20: 427-438. 10.1089 / 10799
12379.
CAS Статья PubMed Google Scholar
Пэган Дж. К., Уайли Ф. Г., Джозеф С., Видберг С., Брайант Н. Дж., Джеймс Д. Э., Стоу Дж. Л.: Синтаксин 4 t-SNARE регулируется во время активации макрофагов для функционирования в мембранном движении и секреции цитокинов. Текущая биология. 2003, 13: 156-160. 10.1016 / S0960-9822 (03) 00006-Х.
Текущая биология. 2003, 13: 156-160. 10.1016 / S0960-9822 (03) 00006-Х.
CAS Статья PubMed Google Scholar
Орлофски А., Вайс Л.М., Кавачи Н., Пристовский М.Б.: Дефицит антиапоптотического белка A1-a приводит к снижению острого воспалительного ответа. J Immunol. 2002, 168: 1840-1846.
CAS Статья PubMed Google Scholar
Wu MX, Ao Z, Prasad KVnS, Wu R, Schlossman SF: IEX-1L, ингибитор апоптоза, участвующий в выживании клеток, опосредованных NF- {kappa} B. Наука. 1998, 281: 998-1001. 10.1126 / science.281.5379.998.
CAS Статья PubMed Google Scholar
De Smaele E, Zazzeroni F, Papa S, Nguyen D, Jin R, Jones J, Cong R, Franzoso G: Индукция gadd45 с помощью NF-B подавляет проапоптотическую передачу сигналов JNK. Природа. 2001, 414: 308-313.10.1038 / 35104560.
CAS Статья PubMed Google Scholar
Jin R, De Smaele E, Zazzeroni F, Nguyen DU, Papa S, Jones J, Cox C, Gelinas C, Franzoso G: Регулирование промотора gadd45beta с помощью NF-kappaB. ДНК Cell Biol. 2002, 21: 491-503. 10.1089 / 104454
CAS Статья PubMed Google Scholar
Derenne S, Monia B, Dean NM, Taylor JK, Rapp MJ, Harousseau JL, Bataille R, Amiot M: антисмысловая стратегия показывает, что Mcl-1, а не Bcl-2 или Bcl-xL, является важным выживанием. белок клеток миеломы человека.Кровь. 2002, 100: 194-199. 10.1182 / blood.V100.1.194.
CAS Статья PubMed Google Scholar
Мацумото Р., Хирасима М., Кита Х., Глейх Г.Дж .: Биологическая активность экалектина: новый фактор, активирующий эозинофилы. J Immunol. 2002, 168: 1961-1967.
CAS Статья PubMed Google Scholar

Hume DA, Underhill DM, Sweet MJ, Ozinsky AO, Liew FY, Aderem A: Макрофаги, постоянно подвергающиеся действию липополисахарида и других агонистов, которые действуют через толл-подобные рецепторы, демонстрируют устойчивое и аддитивное состояние активации.BMC Immunol. 2001, 2: 11-10.1186 / 1471-2172-2-11.
PubMed Central CAS Статья PubMed Google Scholar
Munder M, Eichmann K, Moran JM, Centeno F, Soler G, Modolell M: Th2 / Th3-регулируемая экспрессия изоформ аргиназы в мышиных макрофагах и дендритных клетках. J Immunol. 1999, 163: 3771-3777.
CAS PubMed Google Scholar
Хорн П.Дж., Петерсон К.Л.: Сворачивание хроматина высшего порядка: завершение транскрипции. Наука. 2002, 297: 1824-1827. 10.1126 / science.1074200.
CAS Статья PubMed Google Scholar
Джексон Д. А., Помбо А., Иборра Ф .: Баланс транскрипции: анализ метаболизма ядерной РНК в клетках млекопитающих. FASEB J. 2000, 14: 242-254.
А., Помбо А., Иборра Ф .: Баланс транскрипции: анализ метаболизма ядерной РНК в клетках млекопитающих. FASEB J. 2000, 14: 242-254.
CAS PubMed Google Scholar
Рагхаван А., Огилви Р.Л., Рейли С., Абельсон М.Л., Рагхаван С., Васдевани Дж., Кратвол М., Бохьянен П.Р .: Полногеномный анализ распада мРНК в покоящихся и активированных первичных Т-лимфоцитах человека. Nucl Acids Res. 2002, 30: 5529-5538. 10.1093 / нар / gkf682.
PubMed Central CAS Статья PubMed Google Scholar
Донев Р., Хортон Р., Бек С., Донева Т., Вачева Р., Боуэн В. Р., Шеер Д. Привлечение гетерогенного ядерного рибонуклеопротеина A1 in vivo в область LMP / TAP основного комплекса гистосовместимости.J Biol Chem. 2003, 278: 5214-5226. 10.1074 / jbc.M206621200.
CAS Статья PubMed Google Scholar

Magner WJ, Kazim AL, Stewart C, Romano MA, Catalano G, Grande C, Keizer N, Santaniello F, Tomasi TB: Активация экспрессии гена MHC класса I, II и CD40 ингибиторами гистон-деацетилазы. J Immunol. 2000, 165: 7017-7024.
CAS Статья PubMed Google Scholar
Johnson DR: Локус-специфическая конститутивная и индуцированная цитокинами экспрессия гена HLA класса I. J Immunol. 2003, 170: 1894-1902.
CAS Статья PubMed Google Scholar
Харди М., Санидж Э., Герцог П., Овцарек С. Характеристика и транскрипционный анализ кластера генов рецепторов цитокинов хромосомы 16 мыши. Геном млекопитающих. 2003, 14: 105-118. 10.1007 / s00335-002-2225-0.
CAS Статья PubMed Google Scholar
Вендерфер С.Е., Слэк, Джей П., МакКласки, Скотт Т., Монако. Дж. Дж.: Идентификация 40 генов на контиге размером 1 Мб вокруг кластера генов семейства цитокинов IL-4 на хромосоме мыши 11. Геномика. 2000, 63: 354-373. 10.1006 / geno.1999.6100.
Геномика. 2000, 63: 354-373. 10.1006 / geno.1999.6100.
CAS Статья PubMed Google Scholar
Sweet MJ, Hume DA: Передача сигнала эндотоксина в макрофагах. J Leukoc Biol. 1996, 60: 8-26.
CAS PubMed Google Scholar
Hamilton JA, Whitty G, Kola I, Hertzog P: Эндогенный IFN-альфа-бета подавляет синтез ДНК макрофагов, стимулированный колониестимулирующим фактором (CSF) -1, и опосредует ингибирующие эффекты липополисахарида и TNF-альфа. Журнал иммунологии. 1996, 156: 5223-5227.
Google Scholar
De Maeyer E, De Maeyer-Guignard J, Bailey D: Влияние генотипа мыши на продукцию интерферона. I. Линии, конгенные в локусе If-1. Иммуногенетика.1975, 1: 438-443.
Артикул Google Scholar
De Maeyer E, Jullien P, J DM-G, Demant P: Влияние генотипа мыши на продукцию интерферона. II. Распределение аллелей If-1 среди инбредных штаммов и перенос фенотипа путем пересадки клеток костного мозга. Иммуногенетика. 1975, 2: 151-160.
II. Распределение аллелей If-1 среди инбредных штаммов и перенос фенотипа путем пересадки клеток костного мозга. Иммуногенетика. 1975, 2: 151-160.
Артикул Google Scholar
De Maeyer E, De Maeyer-Guignard J, Hall W., Bailey D: Локус, влияющий на уровни циркулирующего интерферона, индуцированный вирусом опухоли молочной железы мыши.J Gen Virol. 1974, 23: 209-211.
CAS Статья PubMed Google Scholar
Dron M, Meritet JF, Dandoy-Dron F, Meyniel JP, Maury C, Tovey MG: Молекулярное клонирование ADIR, нового интерферон-чувствительного гена, кодирующего белок, связанный с торсинами. Геномика. 2002, 79: 315-325. 10.1006 / geno.2002.6709.
CAS Статья PubMed Google Scholar
Eliopoulos AG, Dumitru CD, Wang C-C, Cho J, Tsichlis PN: Индукция COX-2 с помощью LPS в макрофагах регулируется Tpl2-зависимыми сигналами активации CREB. EMBO J. 2002, 21: 4831-4840. 10.1093 / emboj / cdf478.
EMBO J. 2002, 21: 4831-4840. 10.1093 / emboj / cdf478.
CAS Статья PubMed Google Scholar
Мосманн Т., Червински Х., Бонд М., Гедлин М., Коффман Р. Два типа клона мышиных хелперных Т-клеток. I. Определение в соответствии с профилями активности лимфокинов и секретируемых белков.J Immunol. 1986, 136: 2348-2357.
CAS PubMed Google Scholar
Миллс С.Д., Кинкейд К., Альт Дж. М., Хейлман М. Дж., Хилл А. М.: макрофаги M-1 / M-2 и парадигма Th2 / Th3. Журнал иммунологии. 2000, 164: 6166-6173.
CAS Статья PubMed Google Scholar
Мурата Ю., Шимамура Т., Хамуро Дж .: Поляризация баланса Th2 / Th3 зависит от внутриклеточного окислительно-восстановительного статуса тиоловых макрофагов из-за особой продукции цитокинов.Int Immunol. 2002, 14: 201-212. 10.1093 / intimm / 14. 2.201.
2.201.
CAS Статья PubMed Google Scholar
Beck JA, Lloyd S, Hafezparast M, Lennon-Pierce M, Eppig JT, Festing MFW, Fisher EMC: генеалогии инбредных линий мышей. Природа Генетика. 2000, 24: 23-25. 10.1038 / 71641.
CAS Статья PubMed Google Scholar
Kheradmand F, Werner E, Tremble P, Symons M, Werb Z: Роль Rac1 и кислородных радикалов в экспрессии коллагеназы-1, вызванной изменением формы клеток.Наука. 1998, 280: 898-902. 10.1126 / science.280.5365.898.
CAS Статья PubMed Google Scholar
Sundaresan M, Yu Z-X, Ferrans VJ, Sulciner DJ, Gutkind JS, Irani K, Goldschmidt-Clermont PJ, Finkel T: Регулирование образования активных форм кислорода в фибробластах с помощью Rac1. Биохимический журнал. 1996, 318: 379-382.
PubMed Central CAS Статья PubMed Google Scholar
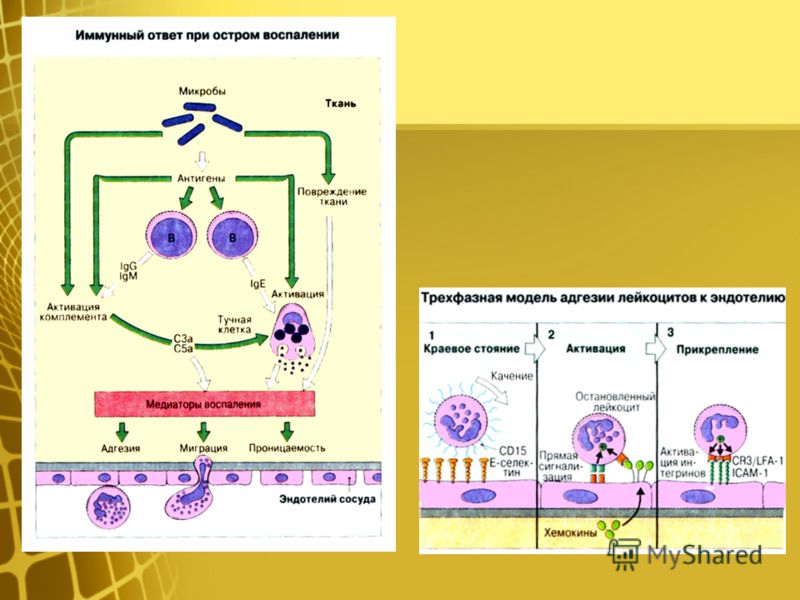
Бартон С., Биггс Т., Бейкер С., Боуэн Н., Аткинсон П.: Nramp1: связь между внутриклеточным транспортом железа и врожденной устойчивостью к внутриклеточным патогенам. J Leukoc Biol. 1999, 66: 757-762.
CAS PubMed Google Scholar
Wyllie S, Seu P, Goss JA: Связанный с естественной резистентностью белок макрофагов 1 Slc11a1 (ранее Nramp1) и метаболизм железа в макрофагах. Микробы и инфекции. 2002, 4: 351-359. 10.1016 / S1286-4579 (02) 01548-4.
CAS Статья PubMed Google Scholar
Себастьяни Г., Олиен Л., Готье С., Скамен Е., Морган К., Грос П., Мало Д. Картирование генетических модуляторов естественной устойчивости к инфекции сальмонеллой тифимуриум у мышей дикого происхождения. Геномика. 1998, 47: 180-186. 10.1006 / geno.1997.5116.
CAS Статья PubMed Google Scholar
Witke W, Sutherland JD, Sharpe A, Arai M, Kwiatkowski DJ: Профилин I необходим для выживания и деления клеток на раннем этапе развития мышей.PNAS. 2001, 98: 3832-3836. 10.1073 / pnas.051515498.
PubMed Central CAS Статья PubMed Google Scholar
Chiang PW, Wang S, Smithivas P, Song WJ, Ramamoorthy S, Hillman J, Puett S, Van Keuren ML, Crombez E, Kumar A: Идентификация и анализ предполагаемого регулятора структуры хроматина человека и мыши SUPT6H andSupt6h. Геномика. 1996, 34: 328-333. 10.1006 / geno.1996.0294.
CAS Статья PubMed Google Scholar
Kjoller L, Kanse SM, Kirkegaard T, Rodenburg KW, Ronne E, Goodman SL, Preissner KT, Ossowski L, Andreasen PA: Ингибитор-1 активатора плазминогена подавляет миграцию клеток, опосредованную интегрином и витронектином, независимо от его функции в качестве ингибитора Активация плазминогена. Экспериментальные исследования клеток. 1997, 232: 420-429. 10.1006 / excr.1997.3540.
Экспериментальные исследования клеток. 1997, 232: 420-429. 10.1006 / excr.1997.3540.
CAS Статья PubMed Google Scholar
May AE, Kanse SM, Lund LR, Gisler RH, Imhof BA, Preissner KT: рецептор урокиназы (CD87) регулирует рекрутирование лейкоцитов через бета 2 интегрины In Vivo.J Exp Med. 1998, 188: 1029-1037. 10.1084 / jem.188.6.1029.
PubMed Central CAS Статья PubMed Google Scholar
Roberts MS, Woods AJ, Shaw PE, Norman JC: ERK1 ассоциирует с интегрином альфа vbeta 3 и регулирует распространение клеток на витронектине. J Biol Chem. 2003, 278: 1975–1985. 10.1074 / jbc.M208607200.
CAS Статья PubMed Google Scholar
Lowenstein C, Alley E, Raval P, Snowman A, Snyder S, Russell S, Murphy W: Ген синтазы оксида азота макрофагов: две вышележащие области опосредуют индукцию интерфероном {гамма} и липополисахаридом. PNAS. 1993, 90: 9730-9734.
PNAS. 1993, 90: 9730-9734.
PubMed Central CAS Статья PubMed Google Scholar
Фогель С.Н., Джонсон Д., Перера П.Я., Медведев А., Ларивьер Л., Куреши С.Т., Мало Д. Передний край: Функциональная характеристика эффекта дефекта C3H / HeJ у мышей, у которых отсутствует ген Lpsn: In Доказательства доминантной отрицательной мутации в естественных условиях.J Immunol. 1999, 162: 5666-5670.
CAS PubMed Google Scholar
Юань Q, Zhao F, Chung S-W, Fan P, Sultzer BM, Kan YW, Wong PMC: Доминирующее отрицательное подавление индуцированного эндотоксином продукции фактора некроза опухоли альфа с помощью Lps (d) / Ran. Труды Национальной академии наук США. 2000, 97: 2852-2857. 10.1073 / pnas.040567797.
CAS Статья Google Scholar
Wong PMC, Yuan Q, Chen H, Sultzer BM, Chung S-W: одноточечная мутация в 3′-нетранслируемой области мРНК Ran приводит к глубоким изменениям в ответах, опосредованных липополисахаридами и эндотоксинами. J Biol Chem. 2001, 276: 33129-33138. 10.1074 / jbc.M105400200.
J Biol Chem. 2001, 276: 33129-33138. 10.1074 / jbc.M105400200.
CAS Статья PubMed Google Scholar
Полторак А., Мерлин Т., Нильсен П.Дж., Сандра О., Смирнова И., Шупп И., Бём Т., Галанос С., Фройденберг М.А.: Точечная мутация в гене IL-12R {beta} 2, лежащем в основе IL- 12 Отсутствие реакции у мышей C57BL / 10ScCr с дефектом Lps.J Immunol. 2001, 167: 2106-2111.
CAS Статья PubMed Google Scholar
Стейси К.Дж., Нагамин Ю.А., Хьюм Д.А. Ингибирование синтеза РНК стабилизирует мРНК урокиназы в макрофагах. FEBS Lett. 1994, 356: 311-313. 10.1016 / 0014-5793 (94) 01294-6.
CAS Статья PubMed Google Scholar
Тушинский Р.Дж., Оливер И.Т., Гильберт Л.Дж., Тайнан П.В., Уорнер-младший, ER.S: Выживание мононуклеарных фагоцитов зависит от клон-специфичного фактора роста, который дифференцированные клетки выборочно уничтожают.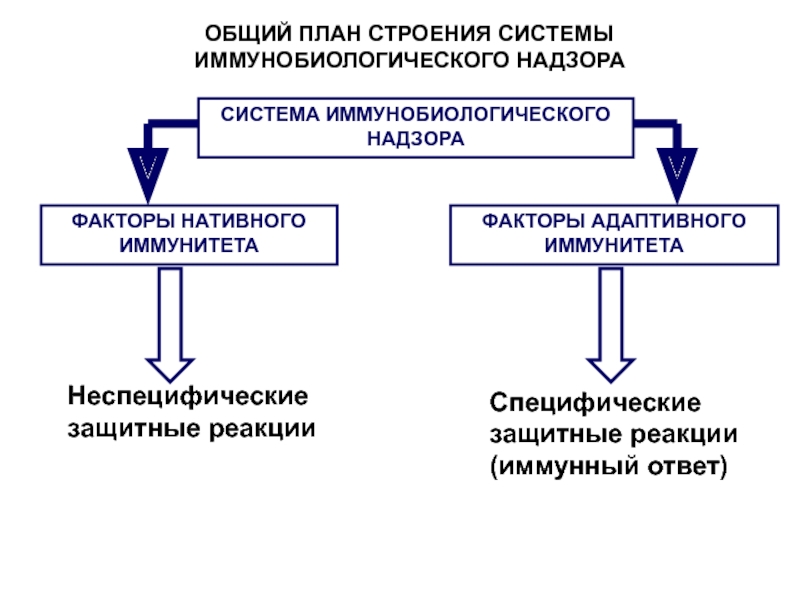 Клетка. 1982, 28: 71-81.
Клетка. 1982, 28: 71-81.
CAS Статья PubMed Google Scholar
Хьюм Д.А., Гордон С. Оптимальные условия для пролиферации макрофагов мышей, полученных из костного мозга, в культуре: роль CSF-1, сыворотки, Ca2 + и приверженность. J. Cell Physiol. 1983, 117: 189-194.
CAS Статья PubMed Google Scholar
Sester DP, Beasley SJ, Sweet MJ, Fowles LF, Cronau SL, Stacey KJ, Hume DA: Бактериальная / CpG ДНК подавляет экспрессию рецептора колониестимулирующего фактора-1 на поверхности макрофагов, полученных из костного мозга мыши, с сопутствующей задержкой роста и фактором -Независимое выживание. Журнал иммунологии. 1999, 163: 6541-6550.
CAS PubMed Google Scholar
Смит GK, Ян YH, Скорость T: Статистические проблемы в анализе данных микрочипа кДНК.В кн .: Функциональная геномика: методы и протоколы.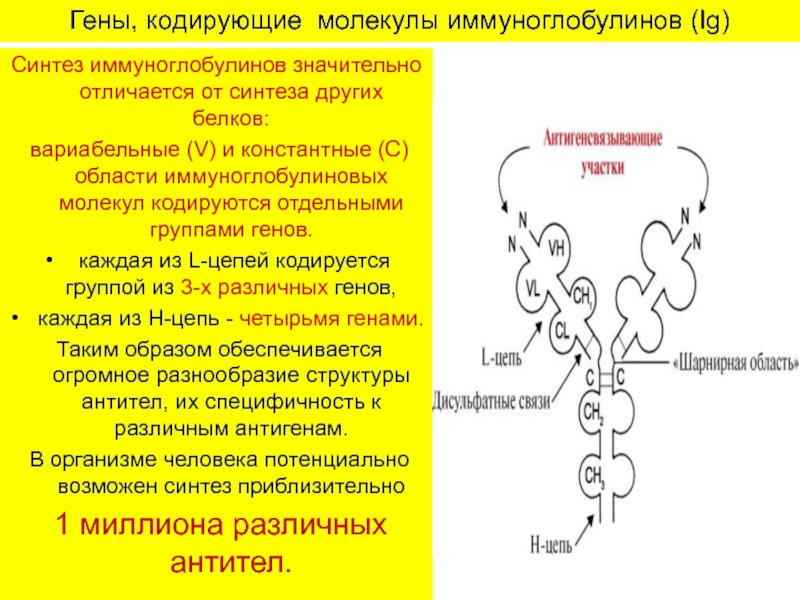 Под редакцией: Браунштейн М.Дж., Ходурский А.Б. 2002, Тотова, Нью-Джерси: Humana Press
Под редакцией: Браунштейн М.Дж., Ходурский А.Б. 2002, Тотова, Нью-Джерси: Humana Press
Google Scholar
Окадзаки Ю., Фуруно М., Касукава Т., Адачи Дж., Боно Х, Кондо С., Никайдо И., Осато Н., Сайто Р., Сузуки Х, Яманака И., Киёсава Х, Яги К., Томару Ю., Хасэгава Ю. , Nogami A, Schonbach C, Gojobori T, Baldarelli R, Hill DP, Bult C, Hume DA, Quackenbush J, Schriml LM, Kanapin A, Matsuda H, Batalov S, Beisel KW, Blake JA, Bradt D, Brusic V, Chothia C, Корбани Л. Е., Казинс С., Далла Е., Драгани Т. А., Флетчер К. Ф., Форрест А., Фрейзер К. С., Гаастерленд Т., Гариболди М., Гисси С., Годзик А., Гоф Дж., Гриммонд С., Густинчич С., Хирокава Н., Джексон И. Дж., Джарвис Э.Д., Канаи А., Кавадзи Х., Кавасава Й., Кедзерски Р.М., Кинг Б.Л., Конагая А., Курочкин IV, Ли И, Ленхард Б., Лайонс, Пенсильвания, Маглотт Д.Р., Мальтийский Л., Маркионни Л., Маккензи Л., Мики Х, Нагашима Т. , Numata K, Okido T, Pavan WJ, Pertea G, Pesole G, Petrovsky N, Pillai R, Pontius JU, Qi D, Ramachandran S, Ravasi T, Reed JC, Reed DJ, Reid J, Ring BZ, Ringwald M, Sandelin A, Schneider C, Semple CA, Setou M, Shimada K, Sultana R, Takenaka Y, Taylor MS, Tea sdale RD, Tomita M, Verardo R, Wagner L, Wahlestedt C, Wang Y, Watanabe Y, Wells C, Wilming LG, Wynshaw-Boris A, Yanagisawa M, et al. : Анализ транскриптома мыши на основе функциональной аннотации 60 770 полноразмерных кДНК. Природа. 2002, 420: 563-573. 10.1038 / природа01266.
: Анализ транскриптома мыши на основе функциональной аннотации 60 770 полноразмерных кДНК. Природа. 2002, 420: 563-573. 10.1038 / природа01266.
Артикул PubMed Google Scholar
Генетический контроль иммунного ответа на инсулин
Розенталь, А.С., Томас, Дж. У., Шроер, Дж., Данхо, В., Бюллесбах, Э. и Фелес, Дж. «Генетический контроль иммунного ответа на инсулин». Инсулин, химический состав, структура и функция инсулина и родственных гормонов: материалы Второго международного симпозиума по инсулину, Ахен, Германия, 4–7 сентября 1979 г. , под редакцией Дитриха Бранденбурга и Акселя Воллмера, Берлин, Бостон: De Gruyter, 2019, стр.559-568. https://doi.org/10.1515/9783111646190-070 Розенталь, А., Томас, Дж., Шроер, Дж., Данхо, В., Бюллесбах, Э. и Фелес, Дж. (2019). Генетический контроль иммунного ответа на инсулин. В D. Brandenburg & A. Wollmer (Ed. ), Инсулин, химия, структура и функция инсулина и родственных гормонов: материалы Второго международного симпозиума по инсулину, Аахен, Германия, 4-7 сентября 1979 г., (стр. 559-568). Берлин, Бостон: Де Грюйтер.https://doi.org/10.1515/9783111646190-070
Розенталь, А., Томас, Дж., Шроер, Дж., Данхо, В., Бюллесбах, Э. и Фелес, Дж. 2019. Генетический контроль иммунного ответа на инсулин. В: Brandenburg, D. and Wollmer, A. ed. Инсулин, химический состав, структура и функция инсулина и родственных гормонов: материалы Второго международного симпозиума по инсулину, Аахен, Германия, 4–7 сентября 1979 г. . Берлин, Бостон: Де Грюйтер, стр. 559-568. https://doi.org/10.1515 / 9783111646190-070
Rosenthal, AS, Thomas, JW, Schroer, J., Danho, W., Büllesbach, E. и Föhles, J .. «Генетический контроль иммунного ответа на инсулин» In Инсулин, химия, структура и функция инсулина и родственные гормоны: Материалы Второго международного симпозиума по инсулину, Ахен, Германия, 4–7 сентября 1979 г.
), Инсулин, химия, структура и функция инсулина и родственных гормонов: материалы Второго международного симпозиума по инсулину, Аахен, Германия, 4-7 сентября 1979 г., (стр. 559-568). Берлин, Бостон: Де Грюйтер.https://doi.org/10.1515/9783111646190-070
Розенталь, А., Томас, Дж., Шроер, Дж., Данхо, В., Бюллесбах, Э. и Фелес, Дж. 2019. Генетический контроль иммунного ответа на инсулин. В: Brandenburg, D. and Wollmer, A. ed. Инсулин, химический состав, структура и функция инсулина и родственных гормонов: материалы Второго международного симпозиума по инсулину, Аахен, Германия, 4–7 сентября 1979 г. . Берлин, Бостон: Де Грюйтер, стр. 559-568. https://doi.org/10.1515 / 9783111646190-070
Rosenthal, AS, Thomas, JW, Schroer, J., Danho, W., Büllesbach, E. и Föhles, J .. «Генетический контроль иммунного ответа на инсулин» In Инсулин, химия, структура и функция инсулина и родственные гормоны: Материалы Второго международного симпозиума по инсулину, Ахен, Германия, 4–7 сентября 1979 г. под редакцией Дитриха Бранденбурга и Акселя Воллмера, 559–568. Берлин, Бостон: De Gruyter, 2019. https://doi.org/10.1515/9783111646190-070.
Розенталь А., Томас Дж., Шроер Дж., Данхо В., Бюллесбах Е., Фёльес Дж.Генетический контроль иммунного ответа на инсулин. В: Brandenburg D, Wollmer A (ed.) Инсулин, химия, структура и функция инсулина и родственных гормонов: материалы Второго международного симпозиума по инсулину, Аахен, Германия, 4–7 сентября 1979 г. . Берлин, Бостон: Де Грюйтер; 2019. С.559-568. https://doi.org/10.1515/9783111646190-070
под редакцией Дитриха Бранденбурга и Акселя Воллмера, 559–568. Берлин, Бостон: De Gruyter, 2019. https://doi.org/10.1515/9783111646190-070.
Розенталь А., Томас Дж., Шроер Дж., Данхо В., Бюллесбах Е., Фёльес Дж.Генетический контроль иммунного ответа на инсулин. В: Brandenburg D, Wollmer A (ed.) Инсулин, химия, структура и функция инсулина и родственных гормонов: материалы Второго международного симпозиума по инсулину, Аахен, Германия, 4–7 сентября 1979 г. . Берлин, Бостон: Де Грюйтер; 2019. С.559-568. https://doi.org/10.1515/9783111646190-070
Генетический контроль иммунного ответа на инфекцию листерией
Цель
Несмотря на неуклонное улучшение здоровья населения, факторы, от централизованного производства продуктов питания до потенциального биотерроризма, сохраняют высокий риск воздействия патогенов пищевого происхождения.В человеческой популяции различия в иммунном ответе на инфекцию определяются уникальными комбинациями аллелей каждого человека.
Наша долгосрочная цель — понять степень вариабельности иммунной функции и определить, как эти вариации влияют на течение и исход инфекции. Listeria monocytogenes — это грамположительная бактерия, которая является одновременно важным патогеном для человека и проверенным инструментом для изучения иммунной функции. Однако трудно изучить генетический контроль чувствительности к L.monocytogenes в человеческих популяциях. С другой стороны, инбредные линии мышей являются генетически поддающимися лечению и демонстрируют широкий диапазон чувствительности к инфекции L monocytogenes.
Для наших исследований генетического контроля восприимчивости к инфекции L. monocytogenes мы выбрали чувствительную, BALB / cByJ и устойчивые линии мышей C57BL / 6ByJ. Наш анализ выживаемости потомства F2 этих штаммов после внутривенного инфицирования L-моноцитогенами выявил два основных локуса количественных признаков (QTL): Listrl на хромосоме 5 и Listr2 на хромосоме 13.
Наша текущая цель — выполнить позиционное клонирование генов, контролирующих дифференциальный ответ на инфекцию L monocytogenes, содержащуюся в этих локусах. Для этого мы: 1. Составим карту локусов восприимчивости L. monocytogenes. Мы проведем обширное генетическое и фенотипическое исследование наших родственных линий мышей, уменьшим размер локусов и определим их границы. 2. Определите гены-кандидаты чувствительности к L. monocytogenes. Полиморфные гены, которые могут контролировать дифференциальную чувствительность к L.monocytogenes будет идентифицироваться комбинацией позиционного клонирования и стратегии, основанной на гипотезах. 3. Определить роль полиморфных генов в контроле инфекции L. monocytogenes.
Для этого мы: 1. Составим карту локусов восприимчивости L. monocytogenes. Мы проведем обширное генетическое и фенотипическое исследование наших родственных линий мышей, уменьшим размер локусов и определим их границы. 2. Определите гены-кандидаты чувствительности к L. monocytogenes. Полиморфные гены, которые могут контролировать дифференциальную чувствительность к L.monocytogenes будет идентифицироваться комбинацией позиционного клонирования и стратегии, основанной на гипотезах. 3. Определить роль полиморфных генов в контроле инфекции L. monocytogenes.
Мы установим решающую роль CXCL11 в контроле инфекции L monocytogenes с использованием трансгенных подходов. Кроме того, мы создадим основу для анализа дополнительных кандидатов.
Обзор его использования в качестве инструмента для выбора устойчивости к болезням
% PDF-1.7
%
1 0 объект
>
эндобдж
2 0 obj
> поток
2017-08-18T10: 15: 23-07: 002017-08-18T10: 15: 23-07: 002017-08-18T10: 15: 23-07: 00Appligent AppendPDF Pro 5. 5uuid: 4dbada41-a686-11b2-0a00- 782dad000000uuid: 4dbb00cc-a686-11b2-0a00-103552fefd7fapplication / pdf
5uuid: 4dbada41-a686-11b2-0a00- 782dad000000uuid: 4dbb00cc-a686-11b2-0a00-103552fefd7fapplication / pdf
 Lff4XHPRDLwA
Lff4XHPRDLwA Гены женской иммунной системы действуют иначе, чем мужские | Центр
В новом исследовании, опубликованном 29 июля в новом журнале Cell Systems , были взяты обычные образцы крови у 12 здоровых добровольцев, чтобы измерить, как определенные гены включаются и выключаются, и как этот показатель варьируется от человека к человеку.Команда Чанга также посмотрела, сколько изменений происходило в разное время у одних и тех же добровольцев. Исследователи изучали исключительно специализированные иммунные клетки, называемые Т-клетками, которые легко выделить с помощью стандартного анализа крови и легко предоставить добровольцам, и которые являются важным компонентом иммунной системы.
Самым сильным предиктором склонности генов к включению и выключению был пол человека.
Одна из целей исследования заключалась в том, чтобы установить базовую меру того, насколько эта активность переключения генов варьируется среди здоровых людей.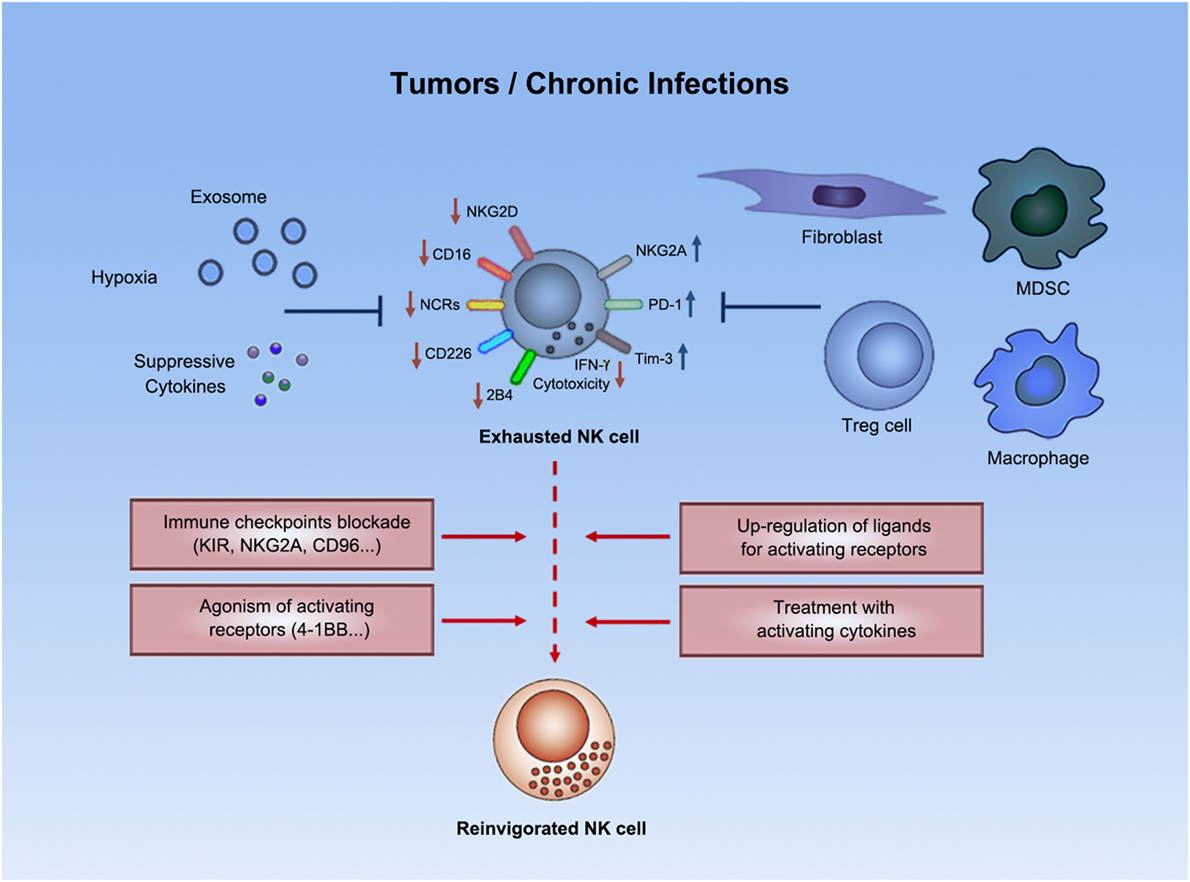 Таким образом, когда другие исследователи проведут аналогичные измерения в отношении больных, они будут иметь представление о том, что является нормальным. Другой целью было усовершенствовать новый метод измерения активности генов в стандартных образцах крови.
Таким образом, когда другие исследователи проведут аналогичные измерения в отношении больных, они будут иметь представление о том, что является нормальным. Другой целью было усовершенствовать новый метод измерения активности генов в стандартных образцах крови.
«Нам было интересно изучить ландшафт регуляции генов непосредственно у живых людей и посмотреть на различия», — сказал Чанг. «Мы спросили:« Насколько люди разные или похожи? »Это отличается от вопроса, имеют ли они одинаковые гены». По его словам, даже у однояйцевых близнецов один из близнецов может иметь аутоиммунное заболевание, а другой — совершенно здоров.И действительно, команда сообщила, что более трети вариаций в активности генов не было связано с генетическими различиями, что предполагает сильную роль окружающей среды. «Я бы сказал, что большая часть различий, вероятно, связана с негенетическим источником», — сказал он.
Половой фактор
У 12 добровольцев 7 процентов генов были включены разными способами от человека к человеку. Для каждого человека эти закономерности сохранялись со временем, как уникальный отпечаток пальца. «Но самым важным предиктором склонности генов к включению и выключению был пол человека.С точки зрения значимости, — сказал Чанг, — секс был намного важнее, чем все остальное, на что мы смотрели, возможно, даже вместе взятые. Когда команда измерила уровни активности генов 30 из 500 лучших генов, которые, как ожидали исследователи, будут проявлять активность, зависящую от пола, они обнаружили, что 20 из 30 генов показали значительную дифференциальную активность между мужчинами и женщинами.
Для каждого человека эти закономерности сохранялись со временем, как уникальный отпечаток пальца. «Но самым важным предиктором склонности генов к включению и выключению был пол человека.С точки зрения значимости, — сказал Чанг, — секс был намного важнее, чем все остальное, на что мы смотрели, возможно, даже вместе взятые. Когда команда измерила уровни активности генов 30 из 500 лучших генов, которые, как ожидали исследователи, будут проявлять активность, зависящую от пола, они обнаружили, что 20 из 30 генов показали значительную дифференциальную активность между мужчинами и женщинами.
Чанг руководит Центром личных динамических регуляторов в Стэнфордском университете, цель которого — составить карту «регуломера» — полного набора всех переключателей, которые включают и выключают гены в реальном времени.
Другими авторами статьи, аффилированными со Стэнфордом, являются Кун Цюй, доктор философии, старший научный сотрудник; Лиза Заба, доктор медицинских наук, инструктор дерматологии; Пол Гиреси, доктор философии, бывший научный сотрудник; Руи Ли, ассистент по исследованиям в области биологических наук; Мишель Лонгмайр, доктор медицины, клинический инструктор по дерматологии; Юн Ким, доктор медицины, Джоан и Питер Хаас младшие, профессор исследования кожных лимфом; и Уильям Гринлиф, доктор философии, доцент генетики.
Эта работа была поддержана Национальным институтом здравоохранения (гранты P50HG007735 и U19AI110491), Медицинским институтом Говарда Хьюза, Стэнфордским онкологическим центром, Фондом исследований склеродермии и Фондом семьи Хааса.
Информация о Департаменте генетики Стэнфорда, который также поддерживал эту работу, доступна на http://genetics.stanford.edu.
Чтобы бороться с инфекциями, нужны нервы: данные о нейроиммунных взаимодействиях, полученные от C. elegans | Модели и механизмы заболеваний
Вдыхание сигаретного дыма связано со значительно повышенным риском микробных инфекций (Nuorti et al., 2000; Obeid and Bercy, 2000). Среди прочего, сигаретный дым подавляет антимикробную активность альвеолярных макрофагов (King et al., 1988). Никотин, активный компонент сигаретного дыма, вызывающий привыкание, вносит важный вклад в пагубное влияние курения на иммунную функцию, поскольку он влияет на центральную и вегетативную нервные системы (Sopori et al. , 1998; Singh et al., 2000). Подобно ACh, никотин связывается с α7 nAChR, подавляя иммунную функцию (Wang et al., 2003). C. elegans обладает богатым набором nAChR (Mongan et al., 1998) и реагирует на никотин аналогично млекопитающим (Feng et al., 2006). Реакции червей на никотин включают острую реакцию при воздействии, реакцию толерантности к хроническому воздействию и реакцию отмены при удалении. Никотиновые ответы в C. elegans также зависят от nAChR (Feng et al., 2006). Недавно был продемонстрирован иммуносупрессивный эффект сигаретного дыма на иммунитет C. elegans (Green et al., 2009). Воздействие сигаретного дыма снижает способность избавляться от инфекций и связано со сниженной экспрессией lpb-7 , который кодирует белок, связывающий жирные кислоты.Интересно, что в эпителиальных клетках бронхов человека, подвергнутых воздействию сигаретного дыма, экспрессия FABP5, гомолога lbp-7 , также снижается (Green et al., 2009), что позволяет предположить, что влияние сигаретного дыма на эпителиальный иммунитет сохраняется.
, 1998; Singh et al., 2000). Подобно ACh, никотин связывается с α7 nAChR, подавляя иммунную функцию (Wang et al., 2003). C. elegans обладает богатым набором nAChR (Mongan et al., 1998) и реагирует на никотин аналогично млекопитающим (Feng et al., 2006). Реакции червей на никотин включают острую реакцию при воздействии, реакцию толерантности к хроническому воздействию и реакцию отмены при удалении. Никотиновые ответы в C. elegans также зависят от nAChR (Feng et al., 2006). Недавно был продемонстрирован иммуносупрессивный эффект сигаретного дыма на иммунитет C. elegans (Green et al., 2009). Воздействие сигаретного дыма снижает способность избавляться от инфекций и связано со сниженной экспрессией lpb-7 , который кодирует белок, связывающий жирные кислоты.Интересно, что в эпителиальных клетках бронхов человека, подвергнутых воздействию сигаретного дыма, экспрессия FABP5, гомолога lbp-7 , также снижается (Green et al., 2009), что позволяет предположить, что влияние сигаретного дыма на эпителиальный иммунитет сохраняется. Опосредуются ли эти эффекты никотином и как белок, связывающий жирные кислоты, опосредует эпителиальный иммунитет, еще предстоит определить. Интересно, что воздействие никотина также увеличивает чувствительность C. elegans к бактериальным инфекциям (Madhumitha Nandakumar and M.-W.T., не опубликовано). Была отмечена связь между курением и подавлением ферментов биосинтеза жирных кислот (в частности, дельта-6-десатуразы, необходимой для биосинтеза жирных кислот омега-3 и омега-6). В C. elegans жирные кислоты омега-3 и омега-6, гамма-линоленовая и стеаридоновая кислоты имеют решающее значение для регуляции врожденного иммунитета. Снижение уровня γ-линоленовой и стеаридоновой кислот снижает активацию p38 MAPK в эпителиальных клетках кишечника и снижает экспрессию антимикробных генов (Nandakumar and Tan, 2008).Влияет ли сигаретный дым на p38 MAPK-опосредованный иммунитет через lbp-7 или через влияние на уровни жирных кислот, остается открытым вопросом. Относительная легкость воздействия на червей токсичных агентов окружающей среды предполагает, что C.
Опосредуются ли эти эффекты никотином и как белок, связывающий жирные кислоты, опосредует эпителиальный иммунитет, еще предстоит определить. Интересно, что воздействие никотина также увеличивает чувствительность C. elegans к бактериальным инфекциям (Madhumitha Nandakumar and M.-W.T., не опубликовано). Была отмечена связь между курением и подавлением ферментов биосинтеза жирных кислот (в частности, дельта-6-десатуразы, необходимой для биосинтеза жирных кислот омега-3 и омега-6). В C. elegans жирные кислоты омега-3 и омега-6, гамма-линоленовая и стеаридоновая кислоты имеют решающее значение для регуляции врожденного иммунитета. Снижение уровня γ-линоленовой и стеаридоновой кислот снижает активацию p38 MAPK в эпителиальных клетках кишечника и снижает экспрессию антимикробных генов (Nandakumar and Tan, 2008).Влияет ли сигаретный дым на p38 MAPK-опосредованный иммунитет через lbp-7 или через влияние на уровни жирных кислот, остается открытым вопросом. Относительная легкость воздействия на червей токсичных агентов окружающей среды предполагает, что C.
