
Ассимиляционная, вентиляционная, запасающая и выделительная ткани
Все эти ткани за исключением некоторых типов выделительной ткани — паренхимные.
Основная паренхима, заполняющая промежутки между сосудисто-волокнистыми пучками и составляющая массу ткани первичной коры и сердцевины, по существу может выполнять все функции, начиная от ассимиляционной и кончая выделительной. Паренхима листьев, входящая в состав так называемого мезофилла (мякоти листа), выполняет преимущественно ассимиляционную функцию, тесно связанную с газообменом, а также с процессом транспирации (парообмена).
На рисунке изображена часть поперечного разреза листа фисташки с очень однородной паренхимной тканью, заключенной между верхним и нижним слоями толстостенного эпидермиса. Это — ассимиляционная ткань.
Ассимиляционная ткань состоит из клеток, расположенных ровными рядами и более или менее сильно вытянутых в направлении, перпендикулярном поверхности органа. Клетки богаты хлоропластами (на рисунке хлоропласты показаны лишь в немногих клетках справа), клеточные оболочки тонкие, клетки довольно тесно соприкасаются друг с другом, межклетники невелики и не всюду имеются.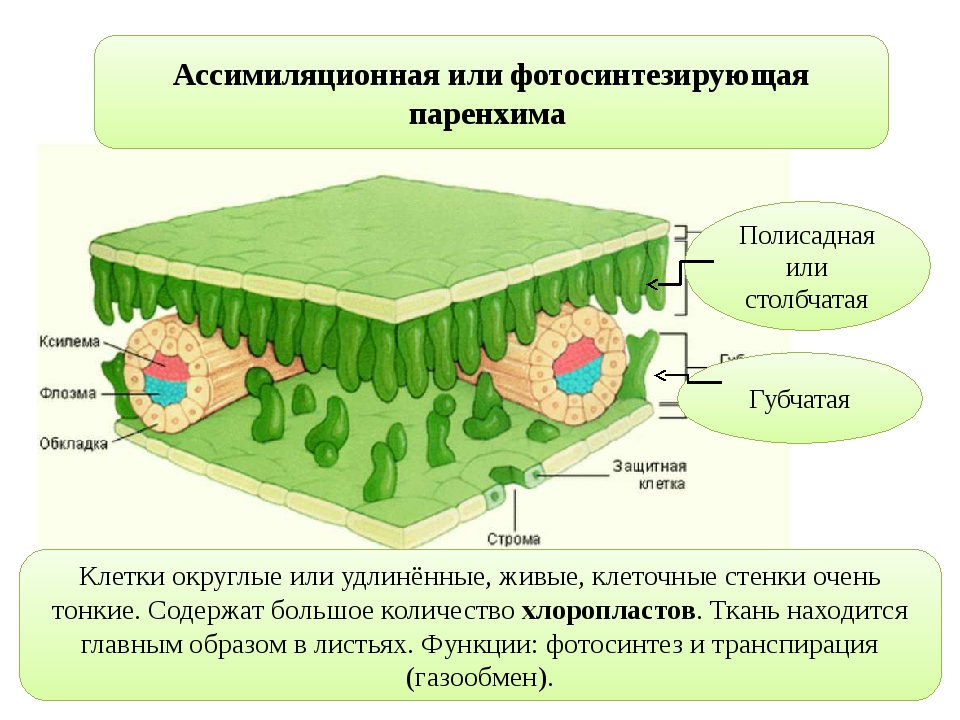
У различных растений столбчатая ассимиляционная ткань построена неодинаково как в отношении длины клеток, их поперечника, так и числа слоев клеток.
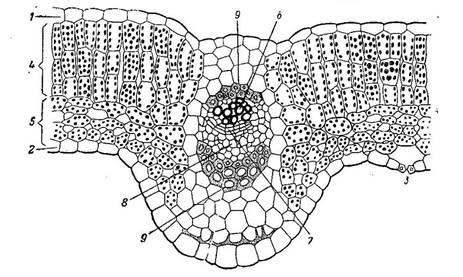
Наряду с тканью, обладающей структурными приспособлениями к ассимиляции, в мезофилле листа очень часто дифференцируется ткань с подчеркнутой специализацией для усиленного газо- и парообмена. На рисунке изображена часть поперечного разреза листа конопли.
Верхняя часть мезофилла состоит из одного слоя столбчатой ткани, а за нею расположена ткань, состоящая из сравнительно мелких клеток весьма разнообразной формы. Самое характерное во второй ткани — большие межклетники. Такую паренхимную ткань в соответствии с ее структурой называют, как указывалось выше, рыхлой, или губчатой, а в соответствии с присущей ей основной специфической функцией ее можно назвать вентиляционной.
В одних случаях, преимущественно у сухопутных растений, воздушные пространства ограничены лишь несколькими соседними клетками, соприкасающимися друг с другом своими отростками. В других случаях и особенно часто у водных растений или обитателей влажных затененных мест воздушные полости весьма обширны и ограничены целым комплексом клеток. Вентиляционная ткань сообщается с наружной средой специальными отверстиями.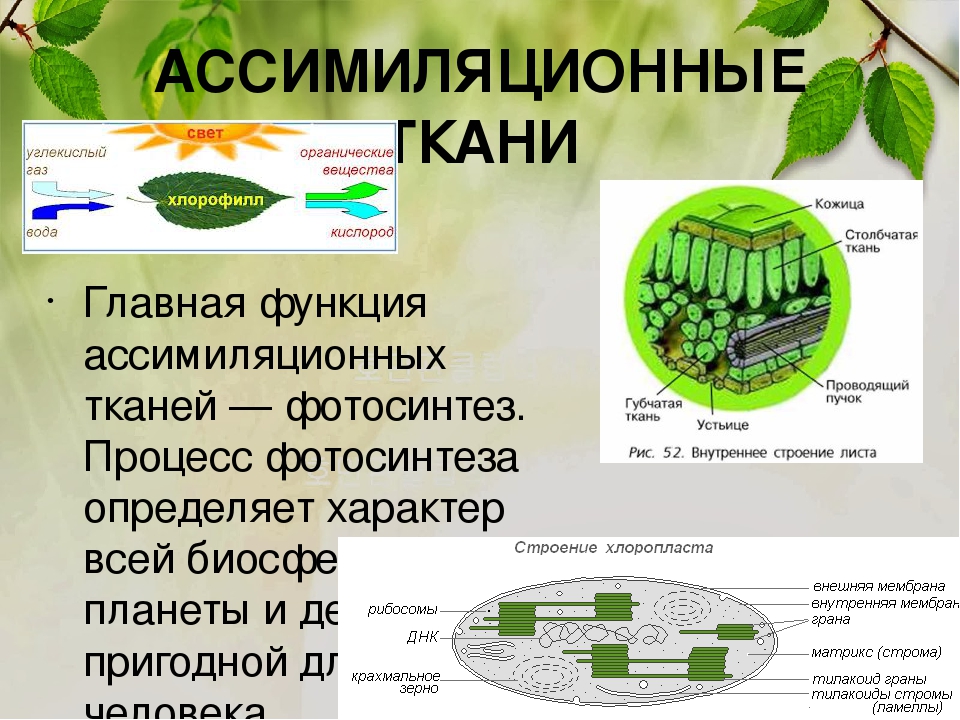 В наиболее распространенной форме выходы вентиляционной ткани представлены устьицами и чечевичками.
В наиболее распространенной форме выходы вентиляционной ткани представлены устьицами и чечевичками.
Характерные примеры запасающей ткани представлены в сердцевине осевых органов, в стебле и корне, а также в плодах и семенах. Кора, во всяком случае в надземных побегах, значительно реже, чем сердцевина, служит для продолжительного хранения запасных веществ. На рисунке изображен фрагмент запасающей ткани из сердцевины фисташки, представляющий собой один из типов запасающей ткани.
Такой ткани свойственны таблитчатая форма клеток и хорошо выраженные простые поры. Толщина клеточных оболочек может быть различной. Запасающая ткань в семенах и плодах чаще всего состоит из клеток с тонкими целлюлозными оболочками, оболочки же клеток сердцевины осевых органов зрелого растения почти всегда одревесневают. Поры в целлюлозных оболочках выявляются с большим трудом, чем поры в одревесневших оболочках, углы не так резки, и вся ткань не имеет того подчеркнуто таблитчатого характера составляющих ее анатомических элементов, как это свойственно клеткам сердцевины.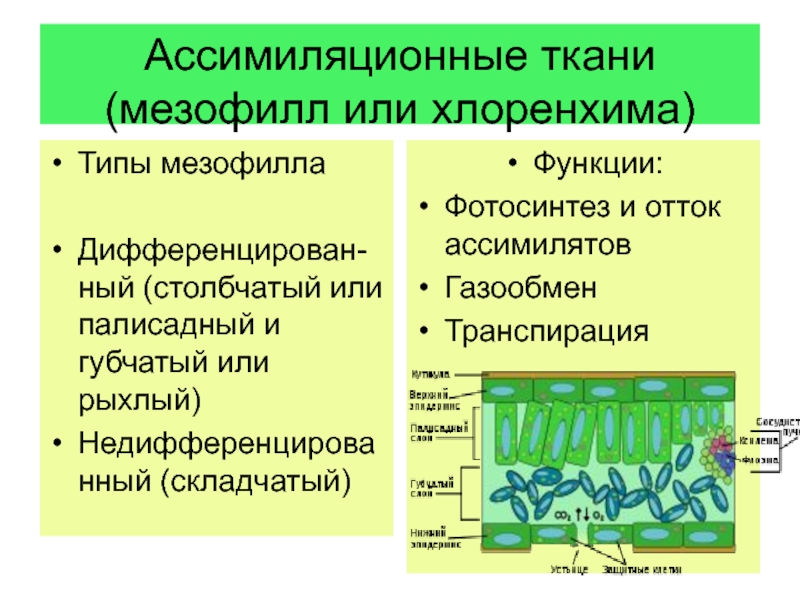
Клетки запасающей ткани плодов и семян накапливают или крахмал, или белок и масла. Крахмал откладывается в виде крахмальных зерен различной величины и формы. Даже в одной и той же клетке, как, например, в клетках эндосперма пшеницы, крахмальные зерна резко различны по величине. 
Выделительная деятельность растений очень разнообразна как в отношении внутренней, так и в отношении наружной секреции. Для первой, с одной стороны, характерно выделение смол в смоляные ходы, а с другой — образование млечного сока в млечниках, возникающих путем разрастания, ветвления и анастомозирования особых гигантских клеток.
Наружные выделительные ткани не столь разнообразны, как внутренние, хотя продукты их выделений не менее многочисленны. Наиболее распространенным типом наружной выделительной ткани являются различные железистые волоски и железки, служащие для выделения эфирных масел, смолистых веществ, нектара и воды. Волоски и железки, выделяющие смолистые вещества, часто встречаются на кроющих чешуях почек и цветков. Эти же выделения придают липкость листьям и стеблям некоторых растений.
Наиболее распространенным типом наружной выделительной ткани являются различные железистые волоски и железки, служащие для выделения эфирных масел, смолистых веществ, нектара и воды. Волоски и железки, выделяющие смолистые вещества, часто встречаются на кроющих чешуях почек и цветков. Эти же выделения придают липкость листьям и стеблям некоторых растений.
Железки, выделяющие нектар, называются нектарниками, а выделяющие воду — гидатодами. Нектарники, как и гидатоды, имеют вид волосков разнообразной формы и строения, размещенных различным образом по поверхности органа, или представляют собой сложную ткань, состоящую из группы клеток, сообщающихся с наружной средой особыми выходами.
Часто выходами для нектара служат устьица, причем устьица либо обычно устроены, либо сильно разрастаются, теряя способность активно регулировать величину своего отверстия.
Устьица последнего типа очень характерны для гидатод. Так же, как и нектарники, гидатоды в качестве приспособления для выделения излишней воды дифференцируются или в виде различных волосков (трихом), или как довольно сложные структурные образования, снабженные специальной тканью для сбора и продвижения воды в капельно-жидком состоянии к устьицу. На рисунке изображена сложная гидатода расширившейся и согнутой ости колоса фуркатного ячменя. Хорошо виден тяж из гидроцитной ткани, идущий от средней жилки к водному устьицу на верхушке изгиба. Для ясности на том же рисунке схематически изображен весь срез расширившейся ости.
На рисунке изображена сложная гидатода расширившейся и согнутой ости колоса фуркатного ячменя. Хорошо виден тяж из гидроцитной ткани, идущий от средней жилки к водному устьицу на верхушке изгиба. Для ясности на том же рисунке схематически изображен весь срез расширившейся ости.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Основные ткани, подготовка к ЕГЭ по биологии
«Грандиозные вещи делаются грандиозными средствами. Одна природа делает великое даром» — Александр И. Герцен
Основные ткани называются так потому, что они составляют основную (бо́льшую) часть массы растения. Им принадлежат важнейшие функции, без которых жизнь
растения совершенно невозможна. В них идет газообмен с окружающей средой, фотосинтез, запасание питательных веществ, запасание воды. Они состоят из живых
паренхиматозных клеток, образованных из первичной меристемы — верхушечной (апикальной).
Начнем изучение с классификации основных тканей.
Ассимиляционная ткань (хлоренхима)
Ассимиляционная — синтезирующая. За счет содержания хлорофилла в данной ткани, здесь активно идет процесс фотосинтеза, хлоропласты в ее клетках выстроены вдоль стенок одним слоем, не затеняя друг друга, подобно солнечным батареям. Наиболее яркий пример местоположения этой ткани — столбчатая ткань мякоти листа (палисадная ткань, от франц. palissade — частокол, загородка), или мезофилл — мягкая ткань, заключенная между двумя слоями эпидермиса в листьях растений.
Хлоренхима расположена непосредственно под эпидермисом, это обеспечивает ее хорошее освещение и газообмен с окружающей средой. Она встречается в надземных органах растений, таких как листья, молодые побеги. Но это не исключает возможность ее возникновения на освещенных корнях, к примеру, в корнях водных растений, воздушных корнях.
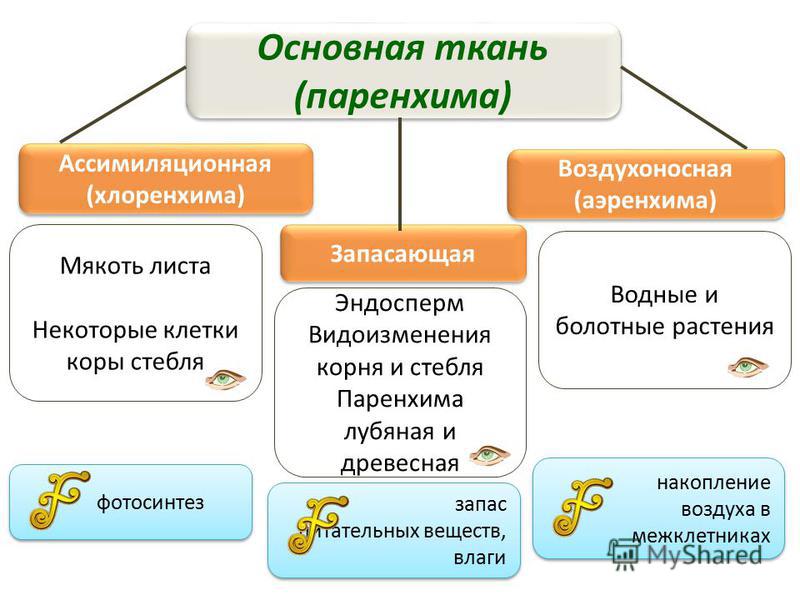
Воздухоносная ткань (аэренхима)
Главная ее функция — газообмен.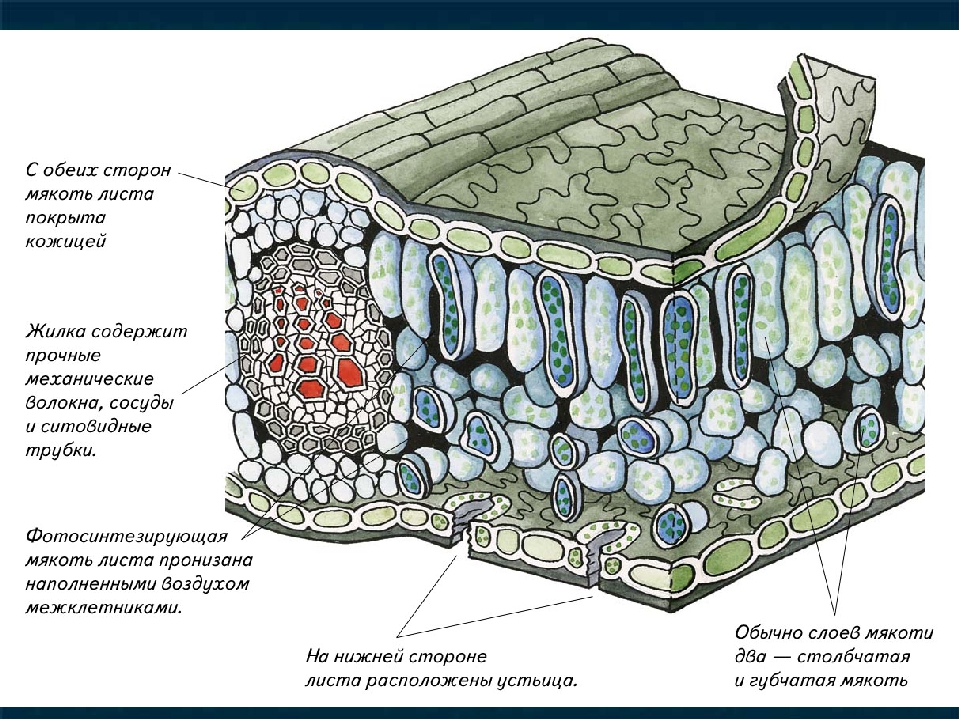 Отличается, прежде всего, наличием межклетников — тканевых пространств, служащих вместилищем для газов. Сквозь устьица воздух
межклетников путем диффузии уравнивается по составу с атмосферным воздухом. В межклетниках из атмосферного воздуха клетки растения поглощают углекислый
газ и выделяют в полость кислород, который затем поступает в окружающую среду.
Отличается, прежде всего, наличием межклетников — тканевых пространств, служащих вместилищем для газов. Сквозь устьица воздух
межклетников путем диффузии уравнивается по составу с атмосферным воздухом. В межклетниках из атмосферного воздуха клетки растения поглощают углекислый
газ и выделяют в полость кислород, который затем поступает в окружающую среду.
Запомните одно из стратегически важных расположений этой ткани — губчатая ткань листа.
У аэренхимы имеется еще одна значимая функция — уменьшение удельного веса растения. Вообразите внутреннюю среду растения, сплошь забитую клеточной массой без всяких промежутков и полостей. Если бы не было аэренхимы, растения, оказавшись тяжелее воды — тонули и опускались на дно, не имея достаточной прочности механической ткани.
Благодаря наличию межклетников в ткани ее удельный вес уменьшается, и она замечательно держится на плаву.
А мы с вами имеем возможность (благодаря аэренхиме! 🙂 получить истинное эстетическое удовольствие от цветущих кувшинок и наслаждаться видом многих других водных растений.
В листьях (на картинке ниже) встречаются клетки с друзой — представляют собой внутриклеточные сростки кристаллов в вакуолях растительных клеток.
Запасающая ткань
Главные функции: запасание и хранение питательных веществ: белков, жиров и углеводов. Преобладает в плодах, сердцевине, луковицах и семенах, клубнях и корневищах. Отдельно отметим, что запасным питательным веществом растений является крахмал.
На рисунке ниже изображен поперечный разрез зоны всасывания корня, видны корневые волоски ризодермы (эпиблемы).
Водоносная паренхима
Клетки этой ткани отличаются большим запасом в вакуолях слизистых веществ, удерживающих влагу. Таким образом, эта ткань способствует удержанию
и запасанию воды. Она хорошо развита у растений, приспособленных к жизни в засушливых местах с сухим климатом. Такие растения получили название —
суккуле́нты от лат. succulentus, «сочный», к ним относятся алоэ, кактусы. Как правило, они произрастают в местах с засушливым климатом.
succulentus, «сочный», к ним относятся алоэ, кактусы. Как правило, они произрастают в местах с засушливым климатом.
Водоносная паренхима при наступлении засухи постепенно отдает свои запасы воды другим, жизненно важным для растения тканям, в первую очередь хлорофиллоносной паренхиме.
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Запасающие и ассимиляционные ткани | Биология. Реферат, доклад, сообщение, краткое содержание, лекция, шпаргалка, конспект, ГДЗ, тест
Тема: Ткани растений
Запасная ткань состоит из паренхимных клеток и большей частью происходит непосредственно из первичной меристемы.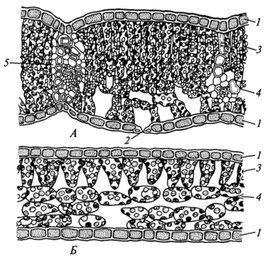 В стебле, например, она находится между перидермой и центральной проводящей частью, а также в сердцевине, сердцевинных лучах и т. д. Среди ткани, образованной паренхимными клетками (основная паренхима), расположены нередко механические ткани, а также и проводящие. Если основная паренхима служит местом накопления питательных веществ, то она называется запасающей тканью.
В стебле, например, она находится между перидермой и центральной проводящей частью, а также в сердцевине, сердцевинных лучах и т. д. Среди ткани, образованной паренхимными клетками (основная паренхима), расположены нередко механические ткани, а также и проводящие. Если основная паренхима служит местом накопления питательных веществ, то она называется запасающей тканью.
В течение вегетационного периода излишки питательных веществ отлагаются в различных частях растения (клубнях, луковицах, древесинной паренхиме стебля и т. д.) в виде запаса на зиму. Паренхимные клетки часто наполнены запасными питательными веществами — крахмалом (например, в клубнях картофеля) или сахаром (например, в корне свеклы), инулином, маслами и т. д. Весной эти вещества идут на развитие листьев, побегов, корней. Материал с сайта http://worldofschool.ru
Ассимиляционная ткань резко выражена в листе, а также и в стебле. Она выполняет одну из важнейших функций зеленого растения — функцию ассимиляции углекислоты воздуха и построения органических веществ. Клетки ассимиляционной ткани паренхиматические и характеризуются наличием в них большого числа хлорофилловых зерен. В листе она образует так называемую мякоть листа. Клетки здесь соединены рыхло, с крупными межклетниками. У большинства растений мякоть листа дифференцирована на 2 слоя клеток. В верхней части пластинки листа клетки удлиненные, прямоугольные, образующие столбчатую, или палисадную, паренхиму. В нижней части — клетки округлые, рыхло соединенные, они образуют губчатую паренхиму. Основная работа по ассимиляции в этом случае протекает в клетках столбчатой паренхимы.
Она выполняет одну из важнейших функций зеленого растения — функцию ассимиляции углекислоты воздуха и построения органических веществ. Клетки ассимиляционной ткани паренхиматические и характеризуются наличием в них большого числа хлорофилловых зерен. В листе она образует так называемую мякоть листа. Клетки здесь соединены рыхло, с крупными межклетниками. У большинства растений мякоть листа дифференцирована на 2 слоя клеток. В верхней части пластинки листа клетки удлиненные, прямоугольные, образующие столбчатую, или палисадную, паренхиму. В нижней части — клетки округлые, рыхло соединенные, они образуют губчатую паренхиму. Основная работа по ассимиляции в этом случае протекает в клетках столбчатой паренхимы.
Ассимиляционные ткани. Строение, функции и размещение.
Ассимиляционные ткани. Строение, функции и размещение.
Ассимиляционная
паренхима, или хлоренхима, характеризуется наличием в ее клетках значительного
количества хлорофилловых зерен. Главной функцией этой ткани является процесс
фотосинтеза, образование его первичных продуктов. Так как образование углеводов
зависит от не только от наличия хлоропластов, но и от солнечного света, то
понятно, что хлоренхима расположена непосредственно под кожицей листьев и
зеленых стеблей, внедряясь иногда и глубже.
Главной функцией этой ткани является процесс
фотосинтеза, образование его первичных продуктов. Так как образование углеводов
зависит от не только от наличия хлоропластов, но и от солнечного света, то
понятно, что хлоренхима расположена непосредственно под кожицей листьев и
зеленых стеблей, внедряясь иногда и глубже.
Хлоренхима
в листьях нередко не образует одной однородной ткани. У многих цветковых
растений она разделяется на две совершенно различных, не связанных между собой
переходами, слоя. Один из них, лежащий под кожицей верхней стороны листа и
состоящий из вытянутых в длину призматических клеток, расположенных
перпендикулярно к поверхности листа, носит название столбчатой, или
палисадной, паренхимы. Клетки ее отличаются присутствием значительного
количества хлоропластов, чем и объясняется интенсивность процесса фотосинтеза в
этой ткани. Другой слой, лежащий с нижней стороны листа, отличается рыхлостью
вследствие образования больших межклеточных пространств, поэтому эта ткань и
называется губчатой паренхимой.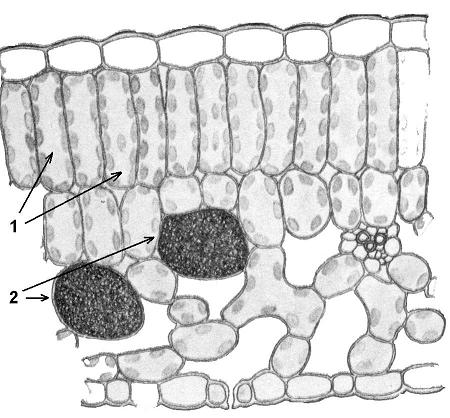
Последняя хотя и имеет в своих клетках хлорофилловые зерна, но ассимиляция является ее второй функцией, а главными функциями являются газообмен и транспирация (испарение воды).
Хлоренхима образуется также в молодых стеблях, органах цветка, плодах и залегает непосредственно под эпидермисом, что обеспечивает её хорошее освещение и газообмен. Хотя к фотосинтезу способны и другие ткани (например, эпидерма), у хлоренхимы эта функция главная и единственная. Обычно фотосинтезирующие ткани располагаются более или менее поверхностно, но иногда хлоренхима располагается в глубине стебля, вокруг пучков или более поверхностно, под механической тканью. В данном случае её функция, вероятно, связана со снабжением внутренних тканей стебля, в первую очередь живых клеток проводящих пучков, кислородом, который образуется в процессе дыхания.
В редких случаях хлоренхима образуется и в корнях, доступных свету, — в корнях водных растений, в воздушных корнях.
Ассимиляционная
ткань чаще всего залегает непосредственно под прозрачной эпидермой.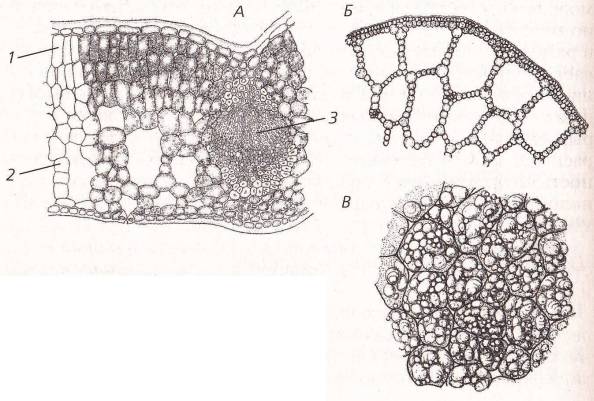 Это
облегчает циркуляцию газов через устьица. Основная масса хлоренхимы
сосредоточена в листьях, меньшая часть — в молодых зеленых стеблях. Нередко в
листьях и стеблях хлоренхима расположена очень рыхло, образуя крупные
газоносные межклетники. В этом случае ассимиляционная функция совмещается с
воздухоносной.
Это
облегчает циркуляцию газов через устьица. Основная масса хлоренхимы
сосредоточена в листьях, меньшая часть — в молодых зеленых стеблях. Нередко в
листьях и стеблях хлоренхима расположена очень рыхло, образуя крупные
газоносные межклетники. В этом случае ассимиляционная функция совмещается с
воздухоносной.
|
Название |
Особенности строения |
Место расположения |
Функции |
|
Покровная 1) кожица 2) пробка 3) кора 4) устьица |
Живые клетки с тонкими и бесцветными оболочками (кожица). Мертвые клетки, одеревеневшие, плотно прилегающие (пробка). Специальные образования- устьица и чечевички (дышат и испаряют). |
Листья, молодые ветки, стволы, корни. Кожица с устьицами. Пробка с чечевичками. |
Защита от неблагоприятных условий. Дыхание и испарение воды. Придают прочность растению. |
|
Проводящая 1) Живые- луб (подкорка) 2) Мертвые (древесина) |
Мертвые клетки в трубках и сосудах, вытянутые в длину. Ситовидные трубки- живые вытянутые клетки без ядра, вакуолей и пластид. |
Корень, стебель, лист, почки, цветки. Лист – стебель- корень. |
Передвижение воды с растворенными в ней веществами (органическими и минеральными). |
|
Механическая 1) Колленхима- вытянутые 2) Паренхима- внутренние 3) Склеренхима- внутренние 4) Каменистые клетки- склереиды |
Живые и мертвые клетки с толстой оболочкой. Каменистые клетки или склереиды- мертвые клетки с одеревеневшими оболочками. Содержатся в плодах, листьях, стеблях, перемешаны с паренхимными клетками, либо существует самостоятельно, без межклеточного вещества. |
Стволы молодых веток, пробка и корки стволов, скорлупа грецкого ореха, покровы семян, мякоть плодов. |
Придают прочность и упругость, способность выдерживать сильную нагрузку. Опора. |
|
Основная
|
Клетки живые, крупные, есть вакуоли, много межклеточного вещества, заполненного млечным соком. В клетках мякоти листа содержатся хлоропласты, участвующие в фотосинтезе.
|
Мякоть листа, мякоть плодов, соцветий, стебля, корней. |
Образование, накопление питательных веществ. Фотосинтез. |
|
Образовательная
|
Живые маленькие клетки с крупными ядрами, без вакуолей. |
Кончик корня, верхушка побега, зародыши, почки, ростковая часть стебля и листа, в коре (между лубом и древесиной). |
Постоянное деление и рост растений, образование тканей. |
|
Секреторная (выделительная) |
Живые тонкостенные клетки. Долгоживущие. Участвуют в метаболизме или запасают вещества в вакуолях. |
Корень, побег, ствол, лист, цветок. |
Вырабатывают, выделяют и сохраняют секреторные продукты (защита, привлечение животных и пр.) |


Ткани растений | Учитель биологии Ольга Коновалова
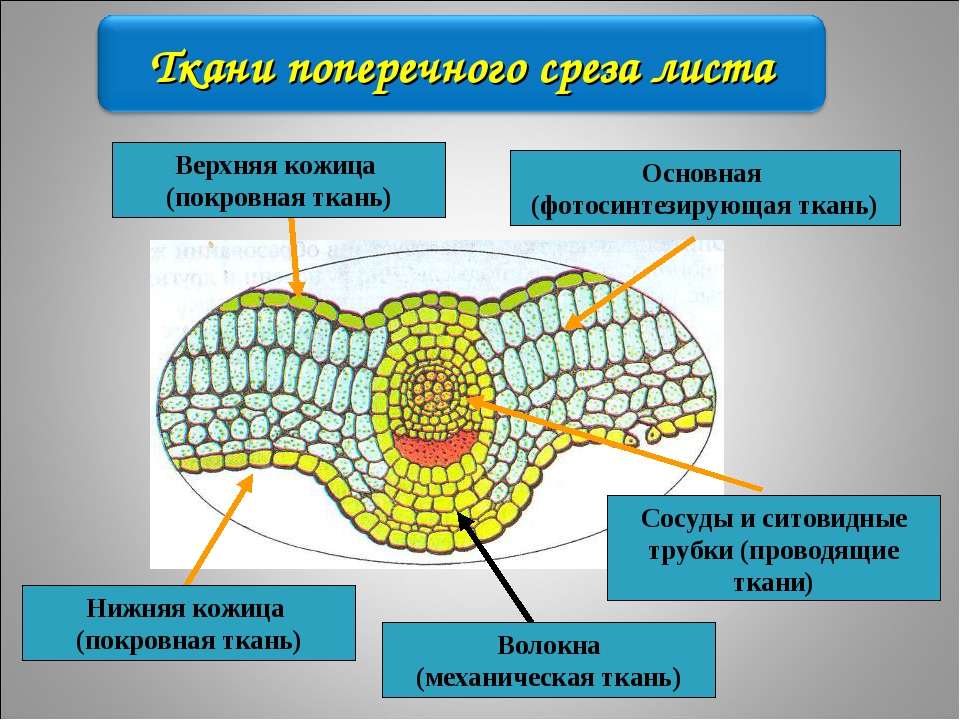
В школьном курсе биологии (в ботанике) рассматривают 5 типов тканей:
1. покровная; 2. механическая; 4. образовательная; 5. проводящая; 3. основная запасающая; 6. основная хлорофиллоносная.
Каждая из этих тканей имеет свои анатомические и физиологические особенности.
| Ткань | Строение и расположение | Значение |
| Покровная | Кожица на поверхности листьев, молодых стеблей, плодах, частях цветка; пробка на поверхности/ Клетки расположены плотно друг к другу, снаружи могут быть покрыты кутикулой (вещество кутин) | Защита от иссушения, защита от механических повреждений, газообмен и транспирация (устьица и чечевички) |
| Механическая | Клетки с сильно утолщеными клеточными стенками:- колленхима состоит из живых сильно вытянутых клеток с неравномерно утолщенными стенками.
— склеренхима состоит из сильно вытянутых мертвых клеток с равномерно одревесневшими стенками, образующих волокна либо склереиды. |
Опорные ткани, обеспечивают прочность, устойчивость |
| Образовательная | Клетки мелкие, быстро делятся митозом. Расположены: верхушечные (конус нарастания и кончик корня), камбий, вставочная (междоузлия стеблей), раневая. | Рост растения в длину, в толщину, восстановление поврежденных тканей, а так же дают начало другим специализированным тканям.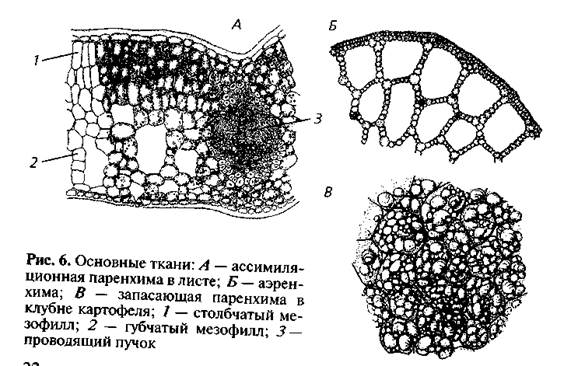 |
| Проводящая | Ксилема (древесина) — мертвые клетки с одревесневшими стенками снабженными порами (трахеиды) и без поперечных перегородок (сосуды).Флоэма (луб) — живые клетки с многочисленными порами в поперечных перегородках (ситовидные трубки) | Ксилема — восходящий ток — вода с минеральными веществами.Флоэма — нисходящий ток — передвижение органических веществ. |
| Основная | Хлорофиллоносная ткань состоит из клеток с большим количеством хлоропластов (в основном в листьях)Запасающая ткань хорошо выражена в стеблях древесных растений, в клубнях, луковицах, плодах, семенах. | Значение хлорофиллоносной ткани — фотосинтез.Соответственно значением является запасание питательных веществ (жиров, белков и углеводов) |
Подробнее можно посмотреть здесь
10 — Лист — СтудИзба
Лекция 10.
Лист
Строение пластинки типичного зеленого листа
Строение нетипичных листьев
Листья злаков
Световые и теневые листья
Листья ксерофитов и суккулентов
Листья тенелюбов
Рекомендуемые файлы
Морфология листьев
Типичный лист представляет собой боковой вырост стебля и состоит из черешка и листовой пластинки. В отличие от стебля и корня, для строения которых характерна радиальная симметрия, в строении листовой пластинки наблюдается симметрия билатеральная, то есть двухсторонняя. В листе имеется верхняя ≈ брюшная и нижняя ≈ спинная сторона (лат. dorsum ≈ спина, venter ≈ брюхо).
В отличие от стебля и корня, для строения которых характерна радиальная симметрия, в строении листовой пластинки наблюдается симметрия билатеральная, то есть двухсторонняя. В листе имеется верхняя ≈ брюшная и нижняя ≈ спинная сторона (лат. dorsum ≈ спина, venter ≈ брюхо).
По своей физиологической роли ≈ лист ≈ орган фотосинтеза, газообмена и транспирации. В тканях листа совершается превращение неорганических веществ (СО2 и Н2О) в органические. Продукты фотосинтеза обычно не накапливаются в листе, а перетекают в другие органы растения.
Основной, типичной категорией листьев являются зеленые, ассимилирующие (питающие) листья, которые называются трофофиллами.
Вместе с тем, листья являются наиболее пластичными, изменчивыми в эволюционном отношении органами. Специализация листа шла и по другим направлениям. Поэтому у растений мы встречаем разнообразнейшие метаморфозы листа, связанные со сменой функций. Но об этом мы поговорим несколько позже.
В связи с указанными функциями в листе хорошо представлены следующие ткани:
| питательная или ассимиляционная; | |
| покровная, регулирующая испарение воды и газообмен; | |
| проводящие такни — обеспечивают подведение почвенных растворов и отток продуктов ассимиляции; | |
| механические ткани, придающие листу прочность. |
Кроме этих четырех тканей, в листе могут встречаться группы клеток или отдельные клетки ≈ идиобласты: склереиды, млечники, места отложения отбросов ≈ минеральных солей и других специфических веществ.
Листья возникают экзогенно (поверхностно), в виде бугорков вблизи от точки роста. Характерно, что в типичном случае листья не имеют верхушечного роста, а нарастают основанием, за счет интеркалярных меристем, причем в течение ограниченного времени.
Эти признаки, однако, не являются абсолютными.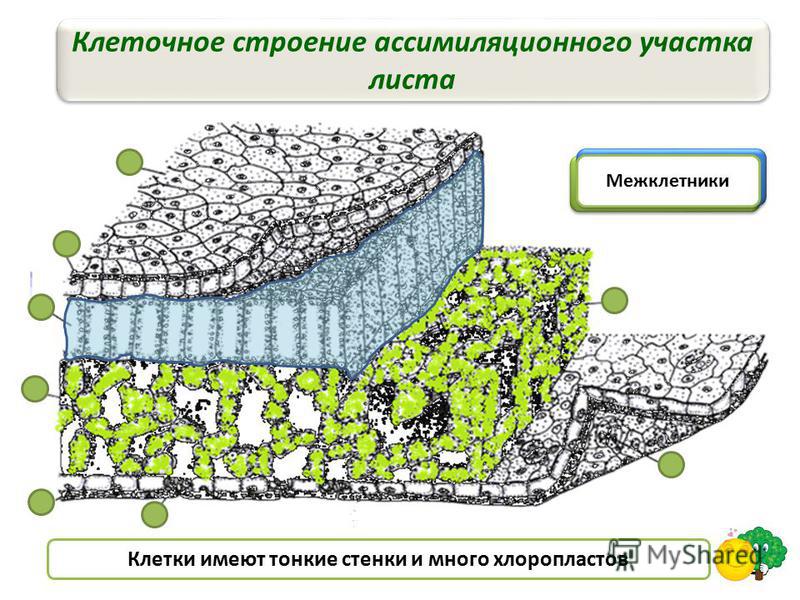
Так, у папоротников, например, долго длится верхушечный рост их крупных листьев ≈ вай.
У некоторых растений листья имеют цилиндрическое строение и радиально-симметричны. Примеры можно найти среди луков и ситников.
Строение пластинки типичного зеленого листа
Лист покрыт со всех сторон эпидермой (кожицей). В обычных, горизонтально ориентированных листьях верхний и нижний эпидермис различаются.
В целом, кожица состоит из тесно сомкнутых паренхимных клеток с извилистыми очертаниями. Клетки нижнего эпидермиса обычно более мелкие и более извилистые.
В верхнем эпидермисе в типичном случае сильнее развиты кутикулярные слои, однако опушение приурочено главным образом к нижней стороне листа. Здесь же, в нижнем эпидермисе обычно располагаются и устьица.
Правда, у водных растений, с плавающими листьями, устьица расположены в верхнем эпидермисе. В верхнем же эпидермисе преобладают устьица у некоторых растений, произрастающих на перегреваемых каменистых склонах.
Типичный эпидермис однослоен. Многослойные варианты связаны обычно с особыми экологическими условиями (как у Ficus).
Под эпидермисом залегает основная зеленая хлорофиллоносная ткань ≈ мезофилл. У большинства растений хлоренхима дифференцирована на столбчатую (палисадную) и рыхлую (губчатую) ткани. В типичном случае к верхней кожице примыкает столбчатый мезофилл, к нижней ≈ губчатый.
Нередко эти две ткани соединяются посредством особых собирательных (воронковидных) клеток.
У многих растений: ветренниц, вейников, бамбуков столбчатая ткань замещается особыми ветвистыми клетками. На границе между палисадной и губчатой тканью располагаются мелкие ответвления проводящих пучков.
Характерным элементом многих листьев являются склереиды, придающие нежным листовым пластинкам дополнительную прочность.
Не менее свойственны листьям крупные клетки ≈ идиобласты с отбросами, например, кристаллами солей (как цистолиты у фикуса).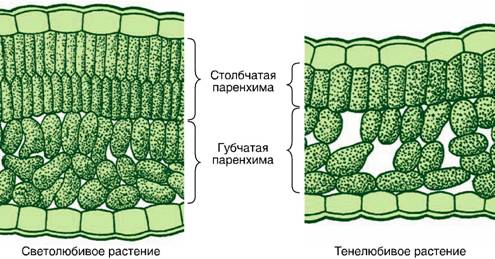 В листьях они имеют особое значение. Так, у некоторых галофитов, растений, произрастающих на засоленных субстратах, единственный способ вывести избыток солей из организма ≈ опадание листьев.
В листьях они имеют особое значение. Так, у некоторых галофитов, растений, произрастающих на засоленных субстратах, единственный способ вывести избыток солей из организма ≈ опадание листьев.
Проводящая система в листе представлена сосудисто-волокнистыми коллатеральными закрытыми пучками. Характерно расположение проводящих тканей: ксилема располагается в верхней части пучков, флоэма ≈ в нижней.
Именно по этому признаку можно точно определить верх и низ листовой пластинки. Все другие признаки изменчивы, так, устьица могут находиться сверху и т. д.
Строение нетипичных листьев
Листья злаков
Особое анатомическое строение имеют листья злаков.
Эпидермис у злаков мощно развит, наружные стенки клеток нередко инкрустируются кремнеземом.
Весьма своеобразны устьица злаков. Замыкающие клетки имеют вид прямоугольников с закругленными концами. Средняя часть каждой из замыкающих клеток очень толстостенна, концевые же участки ≈ тонкостенны.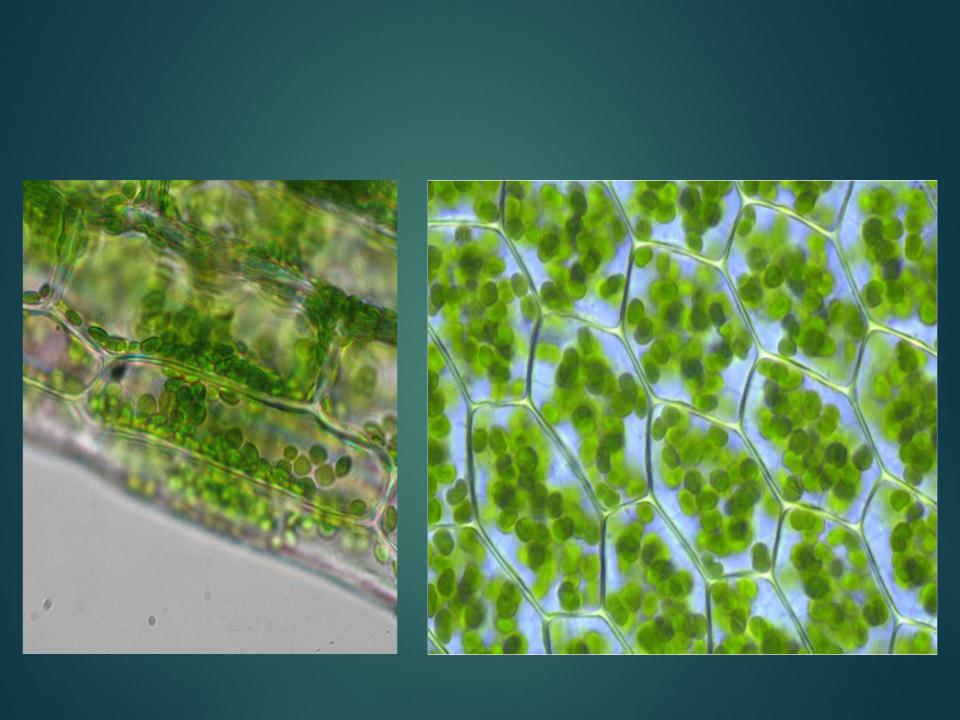 Здесь же имеются пузыревидные вздутия. При повышении тургора они увеличиваются в размерах и устьичная щель раздвигается.
Здесь же имеются пузыревидные вздутия. При повышении тургора они увеличиваются в размерах и устьичная щель раздвигается.
Устьица у злаков обычно располагаются продольными рядами по обеим сторонам листа.
У многих ксерофитных злаков в кожице находится особый аппарат, играющий роль при свертывании и развертывании листьев. Пластинка листа таких злаков сильно ребриста. В ложбинках между ребрами в эпидермисе расположены особые пузыревидные клетки.
Весьма своеобразно у злаков устроен также мезофилл. Здесь нет типичной палисадной и губчатой хлоренхимы. У многих злаков ассимиляционная ткань арсполагается вокруг проводящих пучков в виде обкладки из плотно сомкнутых, удлиненных клеток.
У некоторых злаков клетки ассимиляционной ткани имеют складчатые стенки.
Световые и теневые листья
У деревьев и кустарников листья, находящиеся в условиях более благоприятного освещения, анатомически и морфологически отличаются от листьев, которые сильно затенены.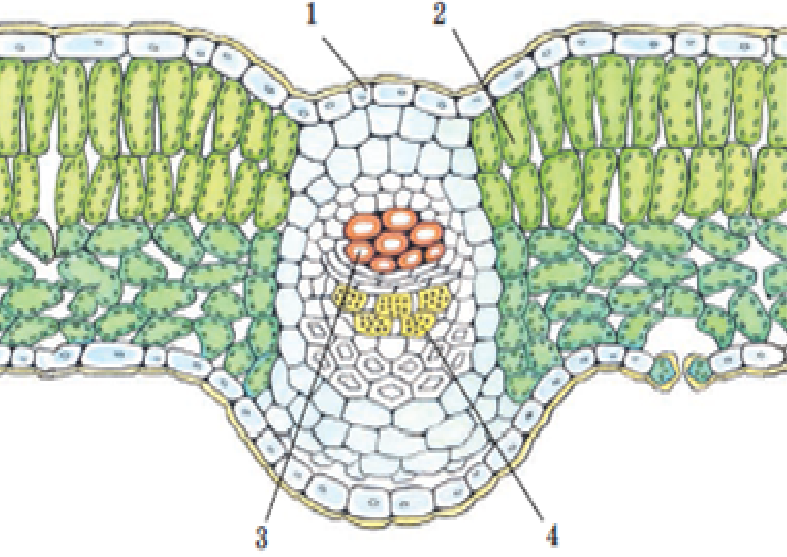
| Световые листья отличаются большей толщиной и жесткостью, кроме того, | |
| клетки кожицы имеют менее волнистые очертания и более толстостенны; | |
| число устьиц на единицу поверхности листа более значительно; | |
| палисадная ткань световых листьев развита мощнее. |
Листья ксерофитов и суккулентов
Весьма своеобразное строение имеют листья растений, приспособившихся переносить длительный засушливый период — ксерофитов, обитающие в условиях дефицита влаги.
Правда, приспособились к этому растения по-разному.
Мясисто-сочные листовые суккуленты: алоэ, толстянки имеют особый водоносный слой в мякоти листа. Они способны быстро накапливать и экономно расходовать влагу.
Примером настоящих ксерофитов являются листья фикуса, панцерий).
Такие растения имеют:
1) очень мощную кутикулу;
2) многослойный эпидермис;
3) нередко на поверхности эпидермиса развиваются мертвые белые ветвистые волоски, хорошо отражающие солнечные лучи и предохраняющие растения от перегрева;
4) устьица обычно глубоко спрятаны в особые устьичные карманы.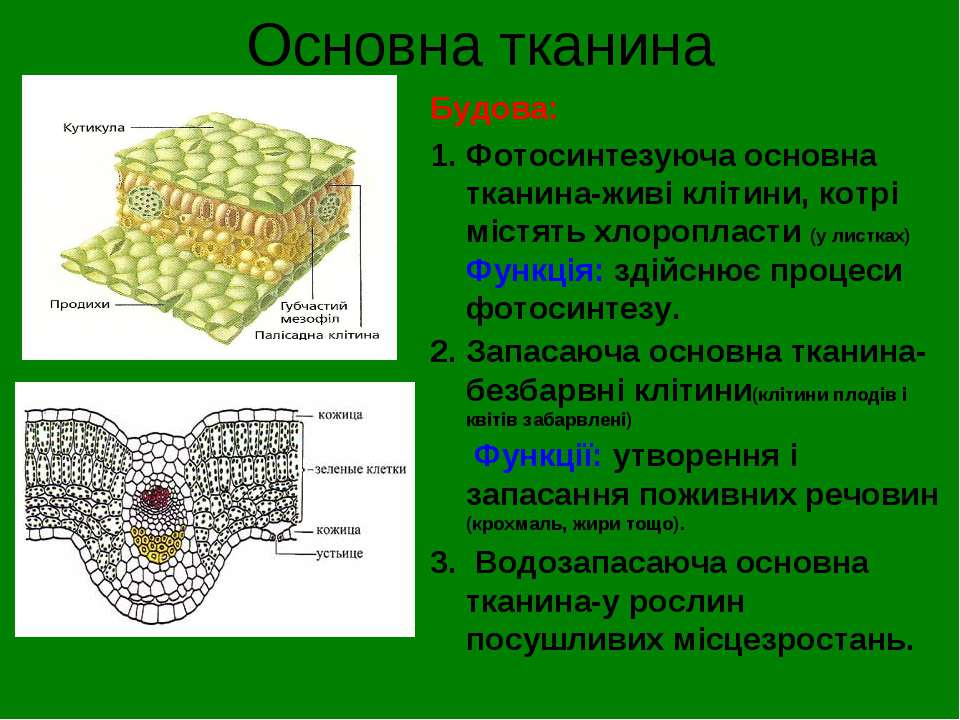
Листья тенелюбов
Особую группу составляют теневые растения (тенелюбы). В эту группу входят многие представители, так называемого, таежного мелкотравья, образующие напочвенный покров в наших темнохвойных лесах: заячья кисличка, адокса мускатная.
Многие теневые растения относятся к группе гигрофитов. Гигрофиты обитают в условиях постоянной влажности воздуха и почвы. Именно такие условия складываются под пологом густого темнохвойного леса.
Структура теневых листьев приспособлена к возможно полному использованию слабого, рассеянного света.
Ассимиляционная ткань имеет малую толщину. У таких растений нередко даже эпидермис богат хлорофиллом. У некоторых тропических бегоний палисадные клетки имеют конусовидную форму и работают как линзы, фокусируя лучи света.
Все эти примеры лишний раз подтверждают огромную пластичность листьев.
Морфология листьев
При морфологическом изучении и описании листьев учитываются следующие признаки:
| характер листорасположения; положение листа на стебле; | |
| общая форма листовой пластинки; | |
| степень рассеченности листовой пластинки; | |
| форма основания листа; | |
| форма верхушки листа; | |
| характер края листа; | |
| Вам также может быть полезна лекция «9 Составляющие ЯПВУ». | типы жилкования листьев. |
Типичный лист делится на цилиндрический черенок и листовую пластинку, подобные листья называются черешковыми. Листья имеют основание, иногда переходящее во влагалище.
Если черешок не выражен, как у некоторых злаков, листья называют сидячими.
Листья делят на простые и сложные. Если у листа одна пластинка ≈ его называют простым. Если на одном черешке несколько пластинок ≈ лист называют сложным.
Более подробно тему «Морфология листа» рассматривают на практических занятиях.
Ботаника онлайн: кожные ткани, паренхима и ассимиляционные ткани
Ботаника онлайн: кожные ткани, паренхима и ассимиляционные ткани — мезофиллБотаника онлайн 1996-2004. Никаких дальнейших обновлений, только исторический документ ботанической науки !
Мезофилл – главный Ассимиляционная ткань листьев
Ассимиляция
ткани — в более широком смысле — все те ткани, которые
образованы клетками, содержащими хлоропласты, и поэтому способны
осуществлять фотосинтез. Они находятся в
все зеленые части растения. Важный аспект фотосинтеза
представляет собой интеграцию углекислого газа в органические соединения. То
полученные продукты суммируются как
уподобляется . Листья высшего
заводы, безусловно, являются наиболее важными производственными центрами — если вы
не учитывать одноклеточные водные водоросли. Листья обычно состоят из трех
различных тканей: мезофилла, эпидермиса и сосудистой
ткани. Мезофилл представляет собой
ткани паренхимы. Это настоящая ассимиляция
ткань.В листьях большинства папоротников и фанерогам организована
в столбчатую паренхиму и губчатую
паренхима.
Они находятся в
все зеленые части растения. Важный аспект фотосинтеза
представляет собой интеграцию углекислого газа в органические соединения. То
полученные продукты суммируются как
уподобляется . Листья высшего
заводы, безусловно, являются наиболее важными производственными центрами — если вы
не учитывать одноклеточные водные водоросли. Листья обычно состоят из трех
различных тканей: мезофилла, эпидермиса и сосудистой
ткани. Мезофилл представляет собой
ткани паренхимы. Это настоящая ассимиляция
ткань.В листьях большинства папоротников и фанерогам организована
в столбчатую паренхиму и губчатую
паренхима.
Поперечный срез Helleborus niger лист. Полутонкий разрез мощностью 3 м.
Стенки клеток окрашиваются калькофторином.
Прежде чем мы перейдем к деталям, необходимо понять, что
термины мезофилл и ассимиляционная ткань не должны использоваться в качестве
синонимы, поскольку существуют листья, лишенные хлоропластов, отличающиеся
организация только немного от зеленых листьев.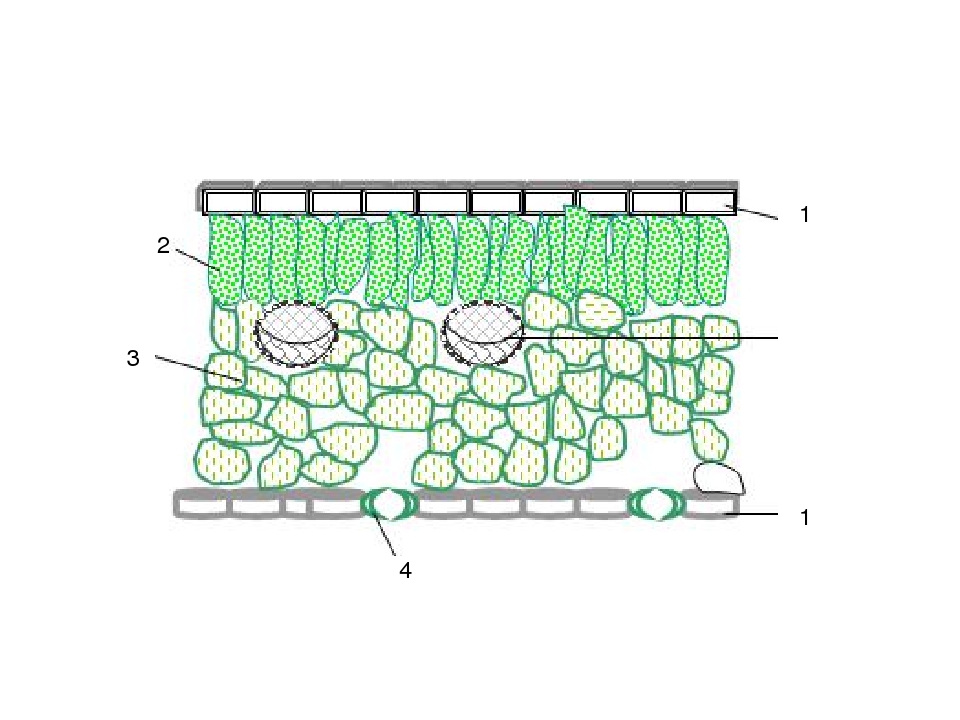 Таким образом, они имеют
мезофилл, но ассимиляции не происходит. Кроме того, ассимиляция
также может встречаться, например, в частях стебля.
Таким образом, они имеют
мезофилл, но ассимиляции не происходит. Кроме того, ассимиляция
также может встречаться, например, в частях стебля.
«Типичный» лист имеет
дорсивентральное строение. То
частокол
паренхима обычно непосредственно
под эпидермисом верхней поверхности листа. То
губчатая паренхима заполняет пространство
под палисадной паренхимой. Он перемежается объемным
межклеточная система, полости которой находятся в непосредственном контакте с
атмосфера через устьица.Это примерно
изложенная структура всегда приводится как главный пример
подбор и адаптация. Наши современные знания о
Фотосинтез показывает, что эффективная ассимиляционная ткань должна
соответствовать следующим критериям:
- Он должен максимально использовать доступный свет.
- Камеры должны постоянно снабжаться достаточным диоксид углерода (CO 2 ).
- Все ячейки должны быть соединены прямым контактом между ячейками с сосудистыми тканями, чтобы гарантировать водоснабжение и транспорт ассимилятов.
структура ассимиляционной ткани листьев выполняет эти требования почти идеальным образом. Кроме того, пластинка тонкая и плоские, а расположение листьев на побегах рассчитано максимально использовать свет. Затенение хлоропластов другими клетками или листьями, таким образом, сводится к минимуму. Поставка растение со светом, углекислым газом и, прежде всего, с водой меняется сильно. Поэтому неудивительно, что модификации этой конструкции с повышенными адаптивными значениями разработали во время эволюции.Их организация зависит от окружающей среды. условия.
Палисад Паренхима
Клетки палисадной паренхимы цилиндрические. Соседний
клетки выглядят как колья частокола. Аналогия поразительна,
при взгляде на поперечное сечение листа, но менее подходит,
когда думаешь о трехмерном расположении клеток.
Клетки палисадной паренхимы содержат в 3–5 раз больше
хлоропласты как у губчатой паренхимы.Хлоропласты остаются
обычно у стенки клетки, так как эта регулировка гарантирует
оптимальное использование света.
Некоторые виды имеют неправильные или даже разветвленные клетки палисадной паренхимы (Y-образные: палисадные клетки). Особенно папоротники, хвойные и некоторые покрытосеменные (некоторые виды Ranunculacea и Caprifoliacea, такие как бузина) содержат такие исключения. Прямых отношений между этими видами не существует. Есть ли разветвленная палисадная паренхима имеет избирательное преимущество? это предположил, что в результате увеличение поверхности выгодно, но опять же: почему эта структура так редка?
Разработка и особенно
на дифференцировку палисадной паренхимы влияют внешние
такие факторы, как свет и содержание CO 2
атмосферы.У многих видов различают солнечно-
и тени листьев. Солнечные листья есть
подвергались воздействию большого количества света в онтогенезе. Этот
приводит к многослойной палисадной паренхиме.
Тень оставляет на контрасте воспринимать
света мало, палисадная паренхима остается однослойной. То
расширение палисадной паренхимы вызывает обычно уменьшение
губчатая паренхима, которая, соответственно, менее развита у
листья солнца. Так же важно, как воздействие света на лист.
положение на стволе.Старые листья, живущие рядом с почвой, часто
имеют палисадную паренхиму толщиной всего в один слой, а
более молодые на верхушке растения обычно имеют многослойные
столбчатые паренхимы. Эта организация наиболее распространена, но
также можно найти следующие варианты:
То
расширение палисадной паренхимы вызывает обычно уменьшение
губчатая паренхима, которая, соответственно, менее развита у
листья солнца. Так же важно, как воздействие света на лист.
положение на стволе.Старые листья, живущие рядом с почвой, часто
имеют палисадную паренхиму толщиной всего в один слой, а
более молодые на верхушке растения обычно имеют многослойные
столбчатые паренхимы. Эта организация наиболее распространена, но
также можно найти следующие варианты:
|
|
Палисадная паренхима на нижней поверхности листа. Это особенно поражает масштабами. Примеры: резюме ( Thuja ), листья черемши ( Allium урсинум ). |
|
|
столбчатая паренхима на обеих поверхностях. Это характерно для
растения сухих местообитаний (ксерофиты). |
|
|
кольцеобразная закрытая палисадная паренхима: встречается в цилиндрически организованные листья и иголки хвойных. |
 Пример: колючий
салат ( Lactuca serriola ).
Пример: колючий
салат ( Lactuca serriola ).
Губчатая Паренхима
Изменчивость клеток губчатого
паренхимы и организация ее даже выше, чем у
палисадная паренхима. Часто говорят, что это
аэренхима , так как она
характеризуется большим количеством сообщающихся межклетников.
Это не означает, что контакт между палисадной паренхимой и
межклеточные пространства менее развиты. Наоборот:
доля палисадной паренхимы, контактирующей с
межклеточное пространство больше, чем у губчатой паренхимы.То
вытянутая и цилиндрическая форма клеток дает возможность только селективного
контакт с соседними клетками даже при плотной упаковке. губчатый
паренхима показывает большие площади контакта, а также межклеточные
контакты между столбчатой паренхимой и губчатой паренхимой и
между губчатой паренхимой и клетками сосудистых пучков. Это
также причина водного и ассимиляционного транспорта, который протекает
без потерь.
губчатый
паренхима показывает большие площади контакта, а также межклеточные
контакты между столбчатой паренхимой и губчатой паренхимой и
между губчатой паренхимой и клетками сосудистых пучков. Это
также причина водного и ассимиляционного транспорта, который протекает
без потерь.
Межклеточная система вызывает развитие большие внутренние поверхности внутри листа.Соотношение межклеточных пространство к общему объему клеток зависит от вида и среды обитания. Это колеблется в пределах 70-700:1000. Ф. М. ТУРРЕЛ получил следующие результаты: при измерении и экстраполяции общей листвы Catalpa в 1934 году:
внутренняя поверхность: 5100 м 2
внешняя поверхность: 390 м 2
Соотношение этих двух поверхностей опять же зависит от видов и естественная среда. Это низкие листья в тени и высокие листья на солнце.
Эволюционные тенденции
Шаг за шагом в ходе эволюции развивались два типа мезофилла. Оба происходят из одной и той же основной ткани и многочисленных переходов
доказывают, что типичные столбчатые и губчатые клетки паренхимы
просто крайности завершенной дифференциации. За немногими исключениями,
все клетки водорослей способны осуществлять фотосинтез. Листья
простые мохообразные, как правило, одно- и лишь изредка
двухслойный. Невозможно провести различие между эпидермисом и
мезофилл.
Оба происходят из одной и той же основной ткани и многочисленных переходов
доказывают, что типичные столбчатые и губчатые клетки паренхимы
просто крайности завершенной дифференциации. За немногими исключениями,
все клетки водорослей способны осуществлять фотосинтез. Листья
простые мохообразные, как правило, одно- и лишь изредка
двухслойный. Невозможно провести различие между эпидермисом и
мезофилл.
Прочее
мохообразные , такие как Polytrichum
-виды (мхи) и Riccia -виды (печеночники)
имеют пластинчатые ребра, состоящие из нескольких слоев ткани поверх
друг друга и приспособлены к ассимиляции.Некоторые печеночники
(как и Marchantia ) имеют многослойное слоевище, которое живет
непосредственно на поверхности почвы. На слое объемистой паренхимы
клетки образуют ассимиляционную ткань из многочисленных мелких, пухлых клеток
собранные в группы и перемежающиеся большими, наполненными воздухом
полости. Печеночники не имеют ни устьиц, ни защитных механизмов. Вместо этого их верхняя поверхность смешана с характерными порами.
что нельзя закрыть.
Вместо этого их верхняя поверхность смешана с характерными порами.
что нельзя закрыть.
Многие водные и болотные растения имеют однородную аэренхиму с очень большими межклеточные полости.Они не структурированы в частокол паренхима и губчатая паренхима. Тоже нельзя отличить между мезофиллом и эпидермисом. Листовая ткань канадской водоросль ( Elodea canadensis ) сокращается до двух клеток слои.
Сосудистые пучки почти всех сосудистых растений окружены связкой оболочка , более или менее обширный слой паренхимы клетки. В его клетках обычно меньше хлоропластов, чем в клетках другие клетки мезофилла. Часто они представляют собой отложения крахмала.
Клетки оболочки пучка Гиса многих, особенно
тропическо-злаковые растения окружены венком из
клетки мезофилла, содержащие хлоропласты (называемые
Анатомия Кранца ; Кранц (немецкий) =
венок). Оба клеточных слоя окружают сосудистые пучки, если они видны на
поперечное сечение в виде двух концентрических колец. Последние годы показали, что
эта конструкция типична для растений C 4 .
C 4 – растения представляют собой группу видов, принадлежащих к разным
одно- и двудольные семейства.Они интегрируют углекислый газ в
различные органические кислоты, прежде чем перейти к фотосинтезу. Растения
растения, у которых отсутствует этот метаболический путь, называются C 3 -растениями.
Даже в пределах одного и того же рода может быть охарактеризован один вид.
как растение C 4 , а другое — как растение C 3 . То
Наиболее известным примером являются два
Atriplex — виды A. patula (C 3 ) и A.
розовая (C 4 ).
Последние годы показали, что
эта конструкция типична для растений C 4 .
C 4 – растения представляют собой группу видов, принадлежащих к разным
одно- и двудольные семейства.Они интегрируют углекислый газ в
различные органические кислоты, прежде чем перейти к фотосинтезу. Растения
растения, у которых отсутствует этот метаболический путь, называются C 3 -растениями.
Даже в пределах одного и того же рода может быть охарактеризован один вид.
как растение C 4 , а другое — как растение C 3 . То
Наиболее известным примером являются два
Atriplex — виды A. patula (C 3 ) и A.
розовая (C 4 ).
Есть даже некоторые виды, у которых нижние листья
построены одним способом, а верхние — другим.Соответственно, у них разная эффективность фотосинтеза. Эти
виды обычно представляют собой суккулентные растения, обитающие в засушливых местообитаниях. То
устьица закрыты в течение дня, чтобы избежать потери воды. Следовательно,
CO 2 можно собирать и накапливать только ночью
(CAM-установки). Фиксация как в C 4 — растения; соответствующий
органическая кислота (обычно малат)
хранится в вакуолях объемистых клеток паренхимы, лишенных
хлоропласты. В течение дня углекислый газ
вновь отщепляется и после диффузии в клетки мезофилла поступает в
пути фотосинтеза.
Следовательно,
CO 2 можно собирать и накапливать только ночью
(CAM-установки). Фиксация как в C 4 — растения; соответствующий
органическая кислота (обычно малат)
хранится в вакуолях объемистых клеток паренхимы, лишенных
хлоропласты. В течение дня углекислый газ
вновь отщепляется и после диффузии в клетки мезофилла поступает в
пути фотосинтеза.
Развитие мезофилла В онтогенезе
Листья развиваются из листьев
контрфорсы , происходящие из небольших боковых утолщений
стрелять. Плоская структура листа обусловлена предпочтительной плоскостью
разделения. Этот тип меристематической активности связан с
пластинчатая меристема ,
предшественник мезофилла. Дифференциация на типичный
столбчатой паренхимы и губчатой паренхимы происходит за счет
неравномерная скорость роста различных слоев клеток в развивающихся
лист.Активность клеточного деления и растяжения клеток происходит в
разные слои в разное время. Кроме того, клетки
расширяться в разные стороны. Пример: клетки
столбчатая паренхима развивается путем антиклинального деления (под прямым углом
к поверхности листа), таковые сосудистой ткани параллельно. В виде
до тех пор, пока деление и удлинение двух тканей согласованы,
соотношение ячеек остается прежним. Но клетки этого
эпидермис начинается довольно рано в онтогенезе, чтобы разделить больше
быстрее, чем у не полностью дифференцированной паренхимы.Этот
вызывает растяжение паренхиматозных клеток, в результате чего межклеточное
образуются пробелы. Они снова частично заполняются при дальнейшем
антиклинальный отдел палисадной паренхимы.
Кроме того, клетки
расширяться в разные стороны. Пример: клетки
столбчатая паренхима развивается путем антиклинального деления (под прямым углом
к поверхности листа), таковые сосудистой ткани параллельно. В виде
до тех пор, пока деление и удлинение двух тканей согласованы,
соотношение ячеек остается прежним. Но клетки этого
эпидермис начинается довольно рано в онтогенезе, чтобы разделить больше
быстрее, чем у не полностью дифференцированной паренхимы.Этот
вызывает растяжение паренхиматозных клеток, в результате чего межклеточное
образуются пробелы. Они снова частично заполняются при дальнейшем
антиклинальный отдел палисадной паренхимы.
Дифференцировка нижнего эпидермиса и
губчатая паренхима скоординирована несколько более вариабельно. В зависимости от
вид представляет собой деление клеток эпидермиса или
губчатая паренхима остановилась первой. Деление клетки и клетка
удлинения временно отделены друг от друга.Разные
скорость роста различных частей развивающегося листа зависит от
волнообразное движение роста. Эти роста
движения были задокументированы в замедленной съемке. Это показывает
впечатляюще, как распространяется рост и расширение листа бука в
интенсивные волнообразные движения (W. KAUSCH, Universität Bonn, 1986;
производство и дистрибьютор: Institut für Wissenschaftlichen
фильм, Геттинген).
Эти роста
движения были задокументированы в замедленной съемке. Это показывает
впечатляюще, как распространяется рост и расширение листа бука в
интенсивные волнообразные движения (W. KAUSCH, Universität Bonn, 1986;
производство и дистрибьютор: Institut für Wissenschaftlichen
фильм, Геттинген).
© Peter v. Sengbusch — Impressum
Кожные ткани, паренхима и ассимиляционные ткани
Ботаника онлайн: Кожные ткани, паренхима и ассимиляционные тканиБотаника онлайн 1996-2004.Никаких дальнейших обновлений, только исторический документ ботанической науки !
Эпидермис является основной кожной тканью
первичные органы растений над землей. Покрывает побег, листья,
цветы, плоды и семена и выполняет несколько функций. Из их
являются защитой от потери воды, от физических и химических
воздействия и от кормления животными, а также участие в
газообмен и секреция. Специальные клетки или устройства для некоторых
эти функции развивались в процессе эволюции.Легкий и механический
раздражители воспринимаются специфическими рецепторами, т.
рецепторов . Стома
комплексы разбросаны по всему эпидермису и действуют как
места газообмена между листом и атмосферой. Комплексы стомы
состоят из нескольких клеток. Две охранные камеры, которые окружают фактический
стома, пора в эпидермисе, контролирует состояние ее открытия. Они
поддерживаются двумя или более дочерними компаниями
ячейки . Кутикула, структура
который секретируется эпидермальными клетками и покрывает их как
непрерывная пленка помогает предотвратить потерю воды.Это также
назначение трихом, эпидермальный
волосы. Но не единственный. Трихомы функционируют как устройства против
животных, как железы, как оружие или водяные пузырьки.
Специальные клетки или устройства для некоторых
эти функции развивались в процессе эволюции.Легкий и механический
раздражители воспринимаются специфическими рецепторами, т.
рецепторов . Стома
комплексы разбросаны по всему эпидермису и действуют как
места газообмена между листом и атмосферой. Комплексы стомы
состоят из нескольких клеток. Две охранные камеры, которые окружают фактический
стома, пора в эпидермисе, контролирует состояние ее открытия. Они
поддерживаются двумя или более дочерними компаниями
ячейки . Кутикула, структура
который секретируется эпидермальными клетками и покрывает их как
непрерывная пленка помогает предотвратить потерю воды.Это также
назначение трихом, эпидермальный
волосы. Но не единственный. Трихомы функционируют как устройства против
животных, как железы, как оружие или водяные пузырьки.
Другой кожной тканью является ризодерма,
который охватывает все подземные части растений. Главной его особенностью являются
корневые волоски. Они улучшают поверхность растения и, следовательно, также
его способность поглощать чрезвычайно. Клетки ризодермы выделяют
слизь, образующая
ризосфера , благодаря чему поглощение
дальше надумано.
Они улучшают поверхность растения и, следовательно, также
его способность поглощать чрезвычайно. Клетки ризодермы выделяют
слизь, образующая
ризосфера , благодаря чему поглощение
дальше надумано.
Более старые побеги и корни в норме прошли вторичную рост. Их первичная кожная ткань заменяется вторичной. кожная ткань, кора . То кора образуется феллогеном, вторичным камбием, и с вкраплениями групп клеток, называемых чечевицами. Чечевицы имеют ту же функцию, что и комплексы устьиц в листьях: они места газообмена.
Наземные ткани составляют основную часть всех
ткани. В них встроены все специализированные клетки.Клетки
основная ткань – паренхима
ячейки . Клетки паренхимы обладают ассимиляционными, запасающими и
ранозаживляющие функции. Паренхима
ткани – это мезофилл,
важная ассимиляционная ткань, частокол
паренхима и губчатая паренхима
листьев. Дальнейшими структурами листа являются сосудистые пучки и их
пучковые оболочки.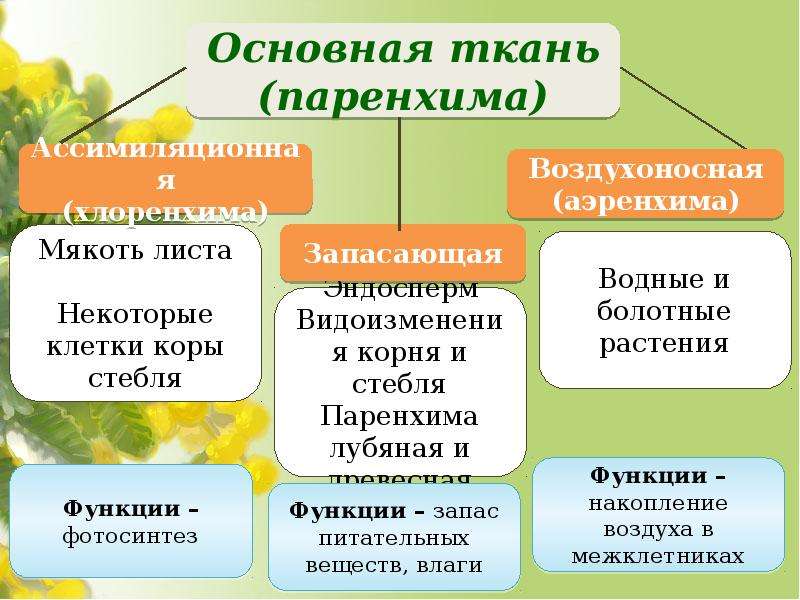 Специализированный способ фиксации углекислого газа
осуществляется венком клеток, который окружает листки пучков
C 4 растений, Kranz
анатомия .Развитие в онтогенезе определяет
форма листа. Его рост происходит волнообразно.
движения.
Специализированный способ фиксации углекислого газа
осуществляется венком клеток, который окружает листки пучков
C 4 растений, Kranz
анатомия .Развитие в онтогенезе определяет
форма листа. Его рост происходит волнообразно.
движения.
© Peter v. Sengbusch — Impressum
Ассимиляция – определение и примеры
Ассимиляция
н.
Акт или процесс ассимиляции; состояние ассимиляции. В биологии это связано с фотосинтезом у автотрофов и процессом пищеварения у гетеротрофов.
Ассимиляция Определение
Что такое ассимиляция? Ассимиляция в биологии определяется как процесс, в котором живые организмы интегрируют питательные вещества из различных внешних ресурсов в свое тело и используют их для удовлетворения энергетических потребностей, необходимых для поддержания жизни.В другом значении ассимиляция относится к поглощению витаминов, минералов и других химических веществ из пищи в желудочно-кишечном тракте.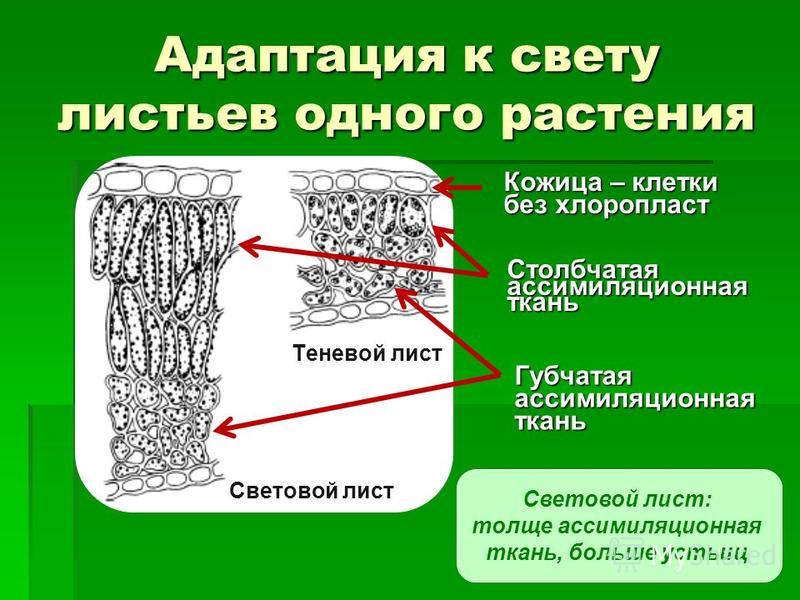 Кроме того, ассимиляция может также относиться к расщеплению сложных молекул, имеющихся в пище, на более простые частицы, а затем к перемещению этих скромных единиц в те места живого тела, где они необходимы, например, к активным клеткам, часто упоминается как . ассимиляция . Процесс фотосинтеза , фиксации азота , преобразования добавок магния и составления биологических тканей и жидкостей посредством адсорбции питательных веществ в организме человека после пищеварения в кишечнике являются некоторыми из практических применений биологических ассимиляция.При этом рост, обновление организмов, структурное развитие, их размножение и восстановление энергетических запасов в организме поддерживаются ассимиляцией. В термодинамике часто упоминается, что человеческие тела представляют собой открытую систему, что означает, что они могут существовать и выживать только при непрерывном поступлении энергий из внешних ресурсов, и, следовательно, это поддерживает значение ассимиляции в биологии.
Кроме того, ассимиляция может также относиться к расщеплению сложных молекул, имеющихся в пище, на более простые частицы, а затем к перемещению этих скромных единиц в те места живого тела, где они необходимы, например, к активным клеткам, часто упоминается как . ассимиляция . Процесс фотосинтеза , фиксации азота , преобразования добавок магния и составления биологических тканей и жидкостей посредством адсорбции питательных веществ в организме человека после пищеварения в кишечнике являются некоторыми из практических применений биологических ассимиляция.При этом рост, обновление организмов, структурное развитие, их размножение и восстановление энергетических запасов в организме поддерживаются ассимиляцией. В термодинамике часто упоминается, что человеческие тела представляют собой открытую систему, что означает, что они могут существовать и выживать только при непрерывном поступлении энергий из внешних ресурсов, и, следовательно, это поддерживает значение ассимиляции в биологии.
Ассимиляция (биологическое определение): превращение пищи в пригодную для использования форму (например,г. жидкое или твердое), которое включается в ткани и органы в результате процессов пищеварения или химического изменения веществ в кровотоке печенью или клеточными секрециями. У растений это относится к процессам фотосинтеза и поглощения сырья, с помощью которых растения получают питание. Примерами ассимиляции являются фотосинтез, фиксация азота и поглощение питательных веществ после переваривания в живую ткань.
Влияние ассимиляции на живые организмы
Солнечное излучение является основным источником энергии для всех живых существ во Вселенной.Живые организмы, обитающие на Земле, в зависимости от использования ими различных форм питания для удовлетворения энергетических потребностей (рис. 1) делятся на два типа:
Организмы, способные производить пищу с помощью света, воды, углекислого газа , и другие химические вещества известны как автотрофные организмы.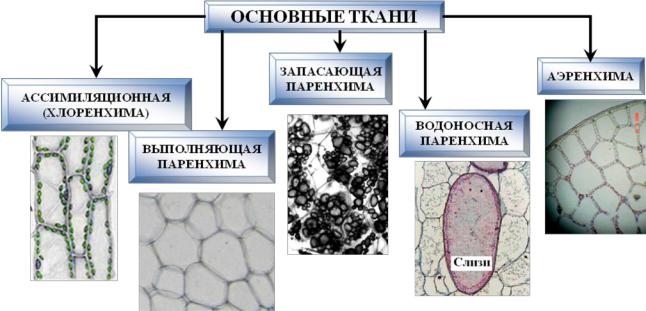 Весь процесс, в котором неорганические соединения в форме различных оксидов углерода превращаются в органические соединения, такие как углеводы, аминокислоты, белки и другие полезные компоненты, называется фиксацией углерода, также называемой ассимиляцией углерода .Автотрофные организмы далее классифицируются на фотоавтотрофы и литоавтотрофы на основе использования солнечного света и процессов неорганического окисления соответственно. С другой стороны, гетеротопные организмы — это те виды существ, которые не способны производить свою собственную пищу, а для получения энергии они полагаются исключительно на получение энергии из внешних ресурсов путем ассимиляции органических веществ, присутствующих в пище, которую они едят. Таким образом, при усвоении сложные белки и углеводы превращаются в глюкозу и аминокислоты.Следовательно, при дыхании используется глюкоза, а аминокислоты усваиваются при построении новых белков. Очень важно понимать, что процессы диссимиляции (восстановления составных частей за счет разрушения) и ассимиляции происходят непрерывно.
Весь процесс, в котором неорганические соединения в форме различных оксидов углерода превращаются в органические соединения, такие как углеводы, аминокислоты, белки и другие полезные компоненты, называется фиксацией углерода, также называемой ассимиляцией углерода .Автотрофные организмы далее классифицируются на фотоавтотрофы и литоавтотрофы на основе использования солнечного света и процессов неорганического окисления соответственно. С другой стороны, гетеротопные организмы — это те виды существ, которые не способны производить свою собственную пищу, а для получения энергии они полагаются исключительно на получение энергии из внешних ресурсов путем ассимиляции органических веществ, присутствующих в пище, которую они едят. Таким образом, при усвоении сложные белки и углеводы превращаются в глюкозу и аминокислоты.Следовательно, при дыхании используется глюкоза, а аминокислоты усваиваются при построении новых белков. Очень важно понимать, что процессы диссимиляции (восстановления составных частей за счет разрушения) и ассимиляции происходят непрерывно.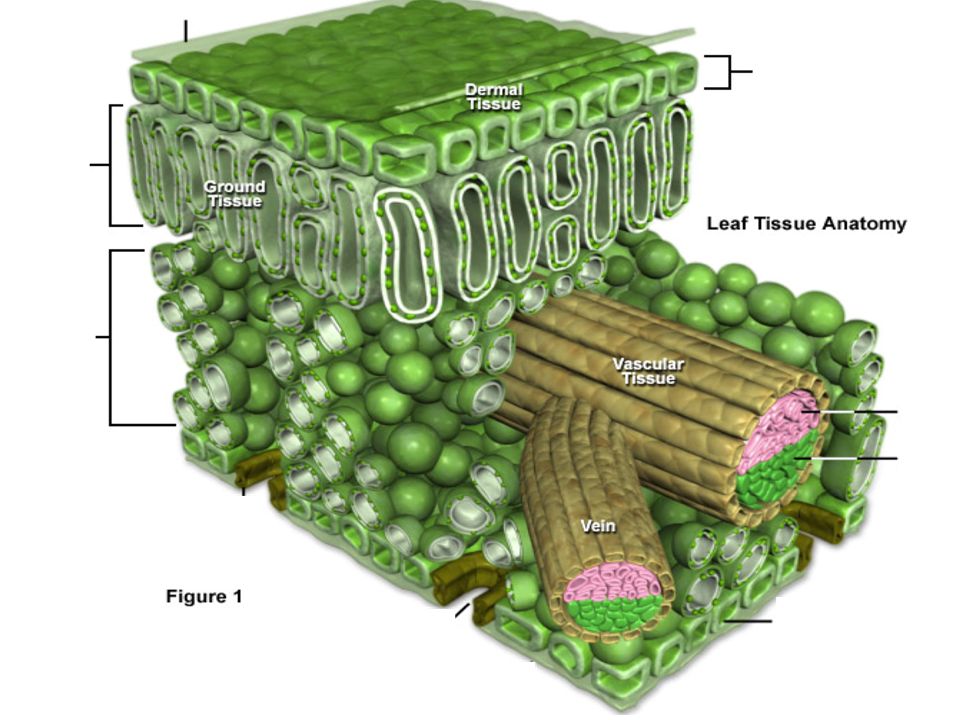 Таким образом, из приведенного выше обсуждения можно сделать вывод, что ассимиляция является очень важным явлением для движения переваренных частиц пищи в различные клетки тела, где требуется энергия.
Таким образом, из приведенного выше обсуждения можно сделать вывод, что ассимиляция является очень важным явлением для движения переваренных частиц пищи в различные клетки тела, где требуется энергия.
Примеры ассимиляции
Что означает ассимиляция с точки зрения процесса? Процесс ассимиляции происходит практически у всех форм живых организмов в мире.Ниже приведены некоторые примеры ассимиляции в различных группах организмов.
Ассимиляция растениями
Что является примером ассимиляции у растений? Азот является основным ингредиентом для выживания и роста растений. Азот поступает к растениям либо с многочисленными удобрениями, либо при поглощении почвой из атмосферы. Процесс ассимиляции в растениях начинается, когда бактерии, присутствующие в почве, превращают азот в аммоний, а затем аммиак снова превращается в нитраты, которые легко усваиваются корнями растений [2].Позже, после цикла поглощения, аминокислоты, нуклеиновые кислоты и хлорофилл строятся вокруг этих поглощенных нитратов. Кроме того, процесс ассимиляции используется и в описанном выше процессе фиксации углерода у автотропных организмов. Следовательно, ассимиляция углерода и ассимиляция азота обнаруживаются у растений и проиллюстрированы на рисунке 2, обозначенными как ассимиляция растениями.
Кроме того, процесс ассимиляции используется и в описанном выше процессе фиксации углерода у автотропных организмов. Следовательно, ассимиляция углерода и ассимиляция азота обнаруживаются у растений и проиллюстрированы на рисунке 2, обозначенными как ассимиляция растениями.
Ассимиляция беспозвоночными
Прием пищи и пищеварение являются двумя наиболее важными процессами ассимиляции у беспозвоночных.Хотя у некоторых беспозвоночных ассимиляция является очень простым процессом, у многих она считается утомительной процедурой. Питательные вещества поглощаются ленточными червями прямо из пищеварительной системы хозяина. Губки и кораллы напрямую поглощают частицы пищи посредством процесса, называемого фагоцитозом. У нематод один длинный пищеварительный тракт, идущий от рта к анусу. Отсюда можно сделать вывод, что все формы беспозвоночных так или иначе берут энергию из питательных веществ, все они используют ассимилированные питательные вещества для роста и получения энергии [4].
Пищеварение человека – ассимиляция
Пищеварительная система человека относительно сложна по сравнению с растениями и насекомыми для усвоения пищи и воды. Пища, введенная в рот, проходит весь путь до желудка, где начинается процесс пищеварения. Инфекционные бактерии уничтожаются соляной кислотой, а более крупные частицы пищи превращаются ассимилированными в более мелкие, которые транспортируются в клетки. Пища после желудка проталкивается в тонкую кишку, где смешивается с печеночной желчью и соками поджелудочной железы.Пища переваривается должным образом, а затем питательные вещества в пище доставляются к различным клеткам тела, где требуется энергия. В дальнейшем после распределения питательных веществ оставшуюся сетку направляют в толстую кишку. Отсюда можно сделать вывод, что процесс усвоения в организме человека происходит в тонкой кишке.
Усвоение питательных веществ
Пища, которую мы едим, усваивается клетками нашего тела. Процесс влечет за собой расщепление пищи на более простые частицы, ее переваривание, а затем распределение по различным частям нашего тела. Более того, как описано ранее, ассимиляция переносит питательные вещества из пищи в клетки, где они используются для роста и размножения. Чтобы понять концепцию усвоения питательных веществ, очень важно пройти цикл переваривания пищи.
Более того, как описано ранее, ассимиляция переносит питательные вещества из пищи в клетки, где они используются для роста и размножения. Чтобы понять концепцию усвоения питательных веществ, очень важно пройти цикл переваривания пищи.
Пищеварительная система в основном начинается изо рта, где пища пережевывается. Слюна, вырабатываемая слюнными железами, смачивает пищу, и поэтому она попадает в желудок через пищевод. Крахмалы также расщепляются слюной на более простые частицы.Когда пища попадает в желудок, очень сильные кислоты и ферменты расщепляют ее на различные питательные вещества, такие как углеводы, белки и жирные кислоты. Следовательно, переваривание пищи частично завершается, а затем она проталкивается в тонкий кишечник, где происходит усвоение питательных веществ. Например, полный цикл переваривания пищи показан на рисунке 3.
Рисунок 3: Полный цикл переваривания пищи в организме человека [5]После частичного переваривания пищи в желудке смесь пищи, жидкостей и пищеварительных соки проходят в тонкой кишке, также называемой чашей, очень регулируемым и контролируемым образом. Тонкая кишка представляет собой туго свернутую трубку, соединяющую желудок и толстую кишку (см. рис. 4).
Тонкая кишка представляет собой туго свернутую трубку, соединяющую желудок и толстую кишку (см. рис. 4). Большая часть процессов пищеварения происходит в тонком кишечнике, и его основной задачей является всасывание и усвоение питательных веществ, содержащихся в пище. У взрослых средняя длина тонкой кишки составляет около 7 метров.
Тонкая кишка была дополнительно разделена на три сегмента: двенадцатиперстную кишку, тощую кишку и подвздошную кишку, как показано на рисунке 4, которые спереди покрыты сальником [6].
Рисунок 4: Анатомические отделы тонкой кишки [7] Каждая часть кишечника играет свою особую роль в абсорбции питательных веществ. На рисунке 4 видно, что пища сначала попадает в двенадцатиперстную кишку, где смешивается с различными выделениями, такими как бикарбонат, пищеварительные ферменты и соли желчных кислот, которые ускоряют процесс пищеварения. Двенадцатиперстная кишка делится на четыре части: нижнюю, верхнюю, восходящую и нисходящую, общей длиной около 25 см. Жиры, содержащиеся в пище, часто перевариваются солями желчи из печени.Углеводы и жиры перевариваются ферментами поджелудочной железы. Кислота, поступающая с пищей из желудка, нейтрализуется бикарбонатом из поджелудочной железы.
Жиры, содержащиеся в пище, часто перевариваются солями желчи из печени.Углеводы и жиры перевариваются ферментами поджелудочной железы. Кислота, поступающая с пищей из желудка, нейтрализуется бикарбонатом из поджелудочной железы.
Правильное переваривание пищи и ее преобразование в питательные вещества связаны со здоровой работой печени и поджелудочной железы. После надлежащего переваривания химуса в двенадцатиперстной кишке он попадает в тощую кишку, где достигается около 90% усвоения питательных веществ, включая белки, углеводы, витамины и минералы.
Тонкая кишка имеет множество мельчайших выступов, называемых микроворсинками. Они используются для увеличения поглощения питательных веществ клетками кишечника. Затем из тощей кишки переваренные питательные вещества высвобождаются в кровоток, чтобы питать и другие клетки.
Наконец, в подвздошной кишке завершается всасывание воды, солей желчных кислот и некоторых избыточных витаминов, после чего лишние частицы пищи перемещаются в толстую или толстую кишку [8].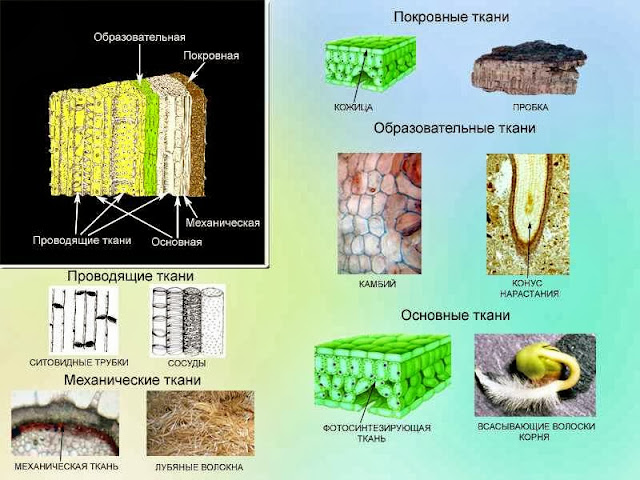
Нарушение всасывания и ассимиляции
Основная роль тонкого кишечника в процессе пищеварения заключается в поглощении питательных веществ из съеденной пищи и доставке их к клеткам через кровоток.Могут быть самые разные синдромы, из-за которых срабатывает плавная работа чаши. Следовательно, эти заболевания называются синдромами мальабсорбции. При нарушении всасывания кишечник не в состоянии всасывать и усваивать как микроэлементы (минералы и витамины), так и макроэлементы (жиры, углеводы и белки). Повреждение кишечника, чрезмерное употребление лекарств, дефицит лактозы, перуксусные заболевания, лучевая терапия и повреждение слизистой оболочки кишечника являются одними из факторов, вызывающих синдром мальабсорбции.Врачи считают, что указанное расстройство можно предсказать по стулу, крови, дыханию, визуализирующим исследованиям и биопсии. Кроме того, нарушение всасывания можно лечить, изменив диету, приняв витаминные и ферментные добавки и отказавшись от молочных продуктов. Считается, что в организме человека из-за синдрома мальабсорбции плохо нарушается адсорбция питательных веществ [9].
Считается, что в организме человека из-за синдрома мальабсорбции плохо нарушается адсорбция питательных веществ [9].
Факторы, влияющие на усвоение
Многие факторы влияют на усвоение питательных веществ. Некоторые из ключевых факторов, которые, по мнению врачей, влияют на усвоение органических веществ, включают температуру, состав потребляемой пищи, временной интервал между двумя приемами пищи, возраст культуры и изменение концентрации и структуры клеток.С другой стороны, исследователи считают, что человеческое тело ведет себя по-разному в процессе усвоения в зависимости от того, какую пищу он принимает. Более того, в различных исследовательских статьях также было опубликовано, что ежедневное употребление зеленого чая и периодическое голодание улучшают систему пищеварения, что приводит к ускоренному процессу усвоения.
Заключение
Из вышеизложенного можно сделать вывод, что распределение питательных веществ между различными клетками является обязательным для их роста, здоровья и размножения. Распространение питательных веществ из тонкой кишки в нужные места осуществляется путем ассимиляции. Считается, что указанный процесс обычно встречается как у автотрофных, так и у гетеротрофных организмов. Более того, в тонкой кишке полный процесс пищеварения происходит в двенадцатиперстной кишке, после чего 90% усвоения питательных веществ, включая белки, углеводы, витамины и минеральные вещества, достигается в тощей кишке. Позже в подвздошной кишке завершается всасывание воды, солей желчных кислот и некоторых избыточных витаминов.Считается, что на процесс усвоения влияют различные факторы, в том числе количество и состав съеденной пищи, временной интервал между двумя приемами пищи, возраст культуры, изменение концентрации и структуры клеток. Кроме того, гладкое выполнение ассимиляции резко нарушается при синдромах мальабсорбции, расстройстве, возникающем из-за неправильной работы тонкой кишки вследствие повреждения в кишечнике, чрезмерного употребления лекарственных средств, лактозной недостаточности, перуксусных заболеваний, лучевой терапии и травм слизистой оболочки кишечника.
Распространение питательных веществ из тонкой кишки в нужные места осуществляется путем ассимиляции. Считается, что указанный процесс обычно встречается как у автотрофных, так и у гетеротрофных организмов. Более того, в тонкой кишке полный процесс пищеварения происходит в двенадцатиперстной кишке, после чего 90% усвоения питательных веществ, включая белки, углеводы, витамины и минеральные вещества, достигается в тощей кишке. Позже в подвздошной кишке завершается всасывание воды, солей желчных кислот и некоторых избыточных витаминов.Считается, что на процесс усвоения влияют различные факторы, в том числе количество и состав съеденной пищи, временной интервал между двумя приемами пищи, возраст культуры, изменение концентрации и структуры клеток. Кроме того, гладкое выполнение ассимиляции резко нарушается при синдромах мальабсорбции, расстройстве, возникающем из-за неправильной работы тонкой кишки вследствие повреждения в кишечнике, чрезмерного употребления лекарственных средств, лактозной недостаточности, перуксусных заболеваний, лучевой терапии и травм слизистой оболочки кишечника.
ПРОЧИТАЙТЕ: Ассимиляция у растений. Учебное пособие по биологии
Попробуйте ответить на приведенный ниже тест, чтобы проверить, что вы уже узнали об ассимиляции (биология).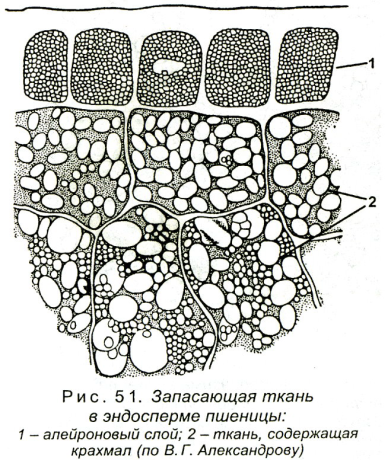
LON-CAPA Ботаника онлайн: кожные ткани, паренхима и ассимиляционные ткани
LON-CAPA Ботаника онлайн: кожные ткани, паренхима и ассимиляционные ткани — мезофилл
Мезофилл – главный Ассимиляционная ткань листьев
Ассимиляция
ткани — в более широком смысле — все те ткани, которые
образованы клетками, содержащими хлоропласты, и поэтому способны
осуществлять фотосинтез.Они находятся в
все зеленые части растения. Важный аспект фотосинтеза
представляет собой интеграцию углекислого газа в органические соединения. То
полученные продукты суммируются как
уподобляется . Листья высшего
заводы, безусловно, являются наиболее важными производственными центрами — если вы
не учитывать одноклеточные водные водоросли. Листья обычно состоят из трех
различных тканей: мезофилла, эпидермиса и сосудистой
ткани. Мезофилл представляет собой
ткани паренхимы. Это настоящая ассимиляция
ткань.В листьях большинства папоротников и фанерогам организована
в столбчатую паренхиму и губчатую
паренхима.
Это настоящая ассимиляция
ткань.В листьях большинства папоротников и фанерогам организована
в столбчатую паренхиму и губчатую
паренхима.
Поперечный срез Helleborus niger лист. Полутонкий разрез мощностью 3 м.
Стенки клеток окрашены калькофторином.
Прежде чем мы перейдем к деталям, необходимо понять, что термины мезофилл и ассимиляционная ткань не должны использоваться в качестве синонимы, поскольку существуют листья, лишенные хлоропластов, отличающиеся организация только немного от зеленых листьев.Таким образом, они имеют мезофилл, но ассимиляции не происходит. Кроме того, ассимиляция также может встречаться, например, в частях стебля.
«Типичный» лист имеет
дорсивентральное строение. То
частокол
паренхима обычно непосредственно
под эпидермисом верхней поверхности листа. То
губчатая паренхима заполняет пространство
под палисадной паренхимой. Он перемежается объемным
межклеточная система, полости которой находятся в непосредственном контакте с
атмосфера через устьица. Это примерно
изложенная структура всегда приводится как главный пример
подбор и адаптация. Наши современные знания о
Фотосинтез показывает, что эффективная ассимиляционная ткань должна
соответствуют следующим критериям:
Это примерно
изложенная структура всегда приводится как главный пример
подбор и адаптация. Наши современные знания о
Фотосинтез показывает, что эффективная ассимиляционная ткань должна
соответствуют следующим критериям:
- Он должен оптимально использовать имеющийся свет.
- Камеры должны постоянно снабжаться достаточным диоксид углерода (CO 2 ).
- Все ячейки должны быть соединены прямым контактом между ячейками с сосудистыми тканями, чтобы гарантировать водоснабжение и транспорт ассимилятов.
структура ассимиляционной ткани листьев выполняет эти
требования почти идеальным образом. Кроме того, пластинка тонкая
и плоские, а расположение листьев на побегах рассчитано
максимально использовать свет. Затенение хлоропластов
другими клетками или листьями, таким образом, сводится к минимуму. Поставка
растение со светом, углекислым газом и, прежде всего, с водой меняется
сильно. Поэтому неудивительно, что модификации
этой конструкции с повышенными адаптивными значениями разработали
во время эволюции.Их организация зависит от окружающей среды.
условия.
Поэтому неудивительно, что модификации
этой конструкции с повышенными адаптивными значениями разработали
во время эволюции.Их организация зависит от окружающей среды.
условия.
Палисад Паренхима
Клетки палисадной паренхимы цилиндрические. Соседний клетки выглядят как колья частокола. Аналогия поразительна, при взгляде на поперечное сечение листа, но менее подходит, когда думаешь о трехмерном расположении клеток. Клетки палисадной паренхимы содержат в 3–5 раз больше хлоропласты как у губчатой паренхимы.Хлоропласты остаются обычно у стенки клетки, так как эта регулировка гарантирует оптимальное использование света.
Некоторые виды имеют неправильные или даже разветвленные
клетки палисадной паренхимы (Y-образные: палисадные
клетки). Особенно папоротники, хвойные и некоторые покрытосеменные (некоторые
виды Ranunculacea и Caprifoliacea, такие как бузина) содержат такие
исключения. Прямых отношений между этими видами не существует. Есть ли
разветвленная палисадная паренхима имеет избирательное преимущество? это
предположил, что в результате увеличение поверхности
выгодно, но опять же: почему эта структура так редка?
Есть ли
разветвленная палисадная паренхима имеет избирательное преимущество? это
предположил, что в результате увеличение поверхности
выгодно, но опять же: почему эта структура так редка?
Разработка и особенно
на дифференцировку палисадной паренхимы влияют внешние
такие факторы, как свет и содержание CO 2
атмосферы.У многих видов различают солнечно-
и тени листьев. Солнечные листья есть
подвергались воздействию большого количества света в онтогенезе. Этот
приводит к многослойной палисадной паренхиме.
Тень оставляет на контрасте воспринимать
света мало, палисадная паренхима остается однослойной. То
расширение палисадной паренхимы вызывает обычно уменьшение
губчатая паренхима, которая, соответственно, менее развита у
листья солнца. Так же важно, как воздействие света на лист.
положение на стволе.Старые листья, живущие рядом с почвой, часто
имеют палисадную паренхиму толщиной всего в один слой, а
более молодые на верхушке растения обычно имеют многослойные
столбчатые паренхимы. Эта организация наиболее распространена, но
также можно найти следующие варианты:
Эта организация наиболее распространена, но
также можно найти следующие варианты:
|
|
Палисадная паренхима на нижней поверхности листа. Это особенно поражает масштабами. Примеры: резюме ( Thuja ), листья черемши ( Allium урсинум ). |
|
|
столбчатая паренхима на обеих поверхностях. Это характерно для растения сухих местообитаний (ксерофиты). Пример: колючий салат ( Lactuca serriola ). |
|
|
кольцеобразная закрытая палисадная паренхима: встречается в цилиндрически организованные листья и иголки хвойных. |
Губчатая Паренхима
Изменчивость клеток губчатого
паренхимы и организация ее даже выше, чем у
палисадная паренхима. Часто говорят, что это
аэренхима , так как она
характеризуется большим количеством сообщающихся межклетников.
Это не означает, что контакт между палисадной паренхимой и
межклеточные пространства менее развиты. Наоборот:
доля палисадной паренхимы, контактирующей с
межклеточное пространство больше, чем у губчатой паренхимы.То
вытянутая и цилиндрическая форма клеток дает возможность только селективного
контакт с соседними клетками даже при плотной упаковке. губчатый
паренхима показывает большие площади контакта, а также межклеточные
контакты между столбчатой паренхимой и губчатой паренхимой и
между губчатой паренхимой и клетками сосудистых пучков. Это
также причина водного и ассимиляционного транспорта, который протекает
без потерь.
Часто говорят, что это
аэренхима , так как она
характеризуется большим количеством сообщающихся межклетников.
Это не означает, что контакт между палисадной паренхимой и
межклеточные пространства менее развиты. Наоборот:
доля палисадной паренхимы, контактирующей с
межклеточное пространство больше, чем у губчатой паренхимы.То
вытянутая и цилиндрическая форма клеток дает возможность только селективного
контакт с соседними клетками даже при плотной упаковке. губчатый
паренхима показывает большие площади контакта, а также межклеточные
контакты между столбчатой паренхимой и губчатой паренхимой и
между губчатой паренхимой и клетками сосудистых пучков. Это
также причина водного и ассимиляционного транспорта, который протекает
без потерь.
Межклеточная система вызывает развитие
большие внутренние поверхности внутри листа.Соотношение межклеточных
пространство к общему объему клеток зависит от вида и среды обитания. Это
колеблется в пределах 70-700:1000.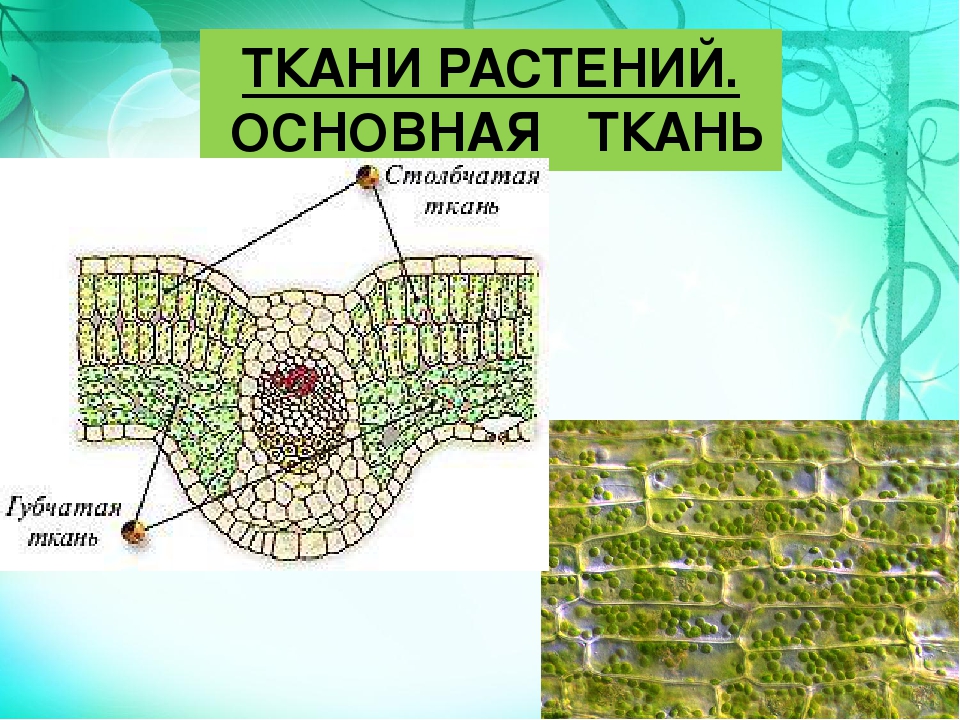 Ф. М. ТУРРЕЛ получил следующие результаты:
при измерении и экстраполяции общей листвы
Catalpa в 1934 году:
Ф. М. ТУРРЕЛ получил следующие результаты:
при измерении и экстраполяции общей листвы
Catalpa в 1934 году:
внутренняя поверхность: 5100 м 2
внешняя поверхность: 390 м 2
Соотношение этих двух поверхностей опять же зависит от видов и естественная среда. Это низкие листья в тени и высокие листья на солнце.
Эволюционные тенденции
Шаг за шагом в ходе эволюции развивались два типа мезофилла.Оба происходят из одной и той же основной ткани и многочисленных переходов доказывают, что типичные столбчатые и губчатые клетки паренхимы просто крайности завершенной дифференциации. За немногими исключениями, все клетки водорослей способны осуществлять фотосинтез. Листья простые мохообразные, как правило, одно- и лишь изредка двухслойный. Невозможно провести различие между эпидермисом и мезофилл.
Прочее
мохообразные , такие как Polytrichum
-виды (мхи) и Riccia -виды (печеночники)
имеют пластинчатые ребра, состоящие из нескольких слоев ткани поверх
друг друга и приспособлены к ассимиляции. Некоторые печеночники
(как и Marchantia ) имеют многослойное слоевище, которое живет
непосредственно на поверхности почвы. На слое объемистой паренхимы
клетки образуют ассимиляционную ткань из многочисленных мелких, пухлых клеток
собранные в группы и перемежающиеся большими, наполненными воздухом
полости. Печеночники не имеют ни устьиц, ни защитных механизмов.
Вместо этого их верхняя поверхность смешана с характерными порами.
что нельзя закрыть.
Некоторые печеночники
(как и Marchantia ) имеют многослойное слоевище, которое живет
непосредственно на поверхности почвы. На слое объемистой паренхимы
клетки образуют ассимиляционную ткань из многочисленных мелких, пухлых клеток
собранные в группы и перемежающиеся большими, наполненными воздухом
полости. Печеночники не имеют ни устьиц, ни защитных механизмов.
Вместо этого их верхняя поверхность смешана с характерными порами.
что нельзя закрыть.
Многие водные и болотные растения имеют однородную аэренхиму с очень большими межклеточные полости.Они не структурированы в частокол паренхима и губчатая паренхима. Тоже нельзя отличить между мезофиллом и эпидермисом. Листовая ткань канадской водоросль ( Elodea canadensis ) сокращается до двух клеток слои.
Сосудистые пучки почти всех сосудистых растений
окружены связкой
оболочка , более или менее обширный слой паренхимы
клетки. В его клетках обычно меньше хлоропластов, чем в клетках
другие клетки мезофилла. Часто они представляют собой отложения крахмала.
Часто они представляют собой отложения крахмала.
Клетки оболочки пучка Гиса многих, особенно
тропическо-злаковые растения окружены венком из
клетки мезофилла, содержащие хлоропласты (называемые
Анатомия Кранца ; Кранц (немецкий) =
венок). Оба клеточных слоя окружают сосудистые пучки, если они видны на
поперечное сечение в виде двух концентрических колец. Последние годы показали, что
эта конструкция типична для растений C 4 .
C 4 – растения представляют собой группу видов, принадлежащих к разным
одно- и двудольные семейства.Они интегрируют углекислый газ в
различные органические кислоты, прежде чем перейти к фотосинтезу. Растения
растения, у которых отсутствует этот метаболический путь, называются C 3 -растениями.
Даже в пределах одного и того же рода может быть охарактеризован один вид.
как растение C 4 , а другое — как растение C 3 . То
Наиболее известным примером являются два
Atriplex — виды A. patula (C 3 ) и A.
розовая (C 4 ).
patula (C 3 ) и A.
розовая (C 4 ).
Есть даже некоторые виды, у которых нижние листья построены одним способом, а верхние — другим.Соответственно, у них разная эффективность фотосинтеза. Эти виды обычно представляют собой суккулентные растения, обитающие в засушливых местообитаниях. То устьица закрыты в течение дня, чтобы избежать потери воды. Следовательно, CO 2 можно собирать и накапливать только ночью (CAM-установки). Фиксация как в C 4 — растения; соответствующий органическая кислота (обычно малат) хранится в вакуолях объемистых клеток паренхимы, лишенных хлоропласты. В течение дня углекислый газ вновь отщепляется и после диффузии в клетки мезофилла поступает в пути фотосинтеза.
Развитие мезофилла В онтогенезе
Листья развиваются из листьев
контрфорсы , происходящие из небольших боковых утолщений
стрелять. Плоская структура листа обусловлена предпочтительной плоскостью
разделения. Этот тип меристематической активности связан с
пластинчатая меристема ,
предшественник мезофилла. Дифференциация на типичный
столбчатой паренхимы и губчатой паренхимы происходит за счет
неравномерная скорость роста различных слоев клеток в развивающихся
лист.Активность клеточного деления и растяжения клеток происходит в
разные слои в разное время. Кроме того, клетки
расширяться в разные стороны. Пример: клетки
столбчатая паренхима развивается путем антиклинального деления (под прямым углом
к поверхности листа), таковые сосудистой ткани параллельно. В виде
до тех пор, пока деление и удлинение двух тканей согласованы,
соотношение ячеек остается прежним. Но клетки этого
эпидермис начинается довольно рано в онтогенезе, чтобы разделить больше
быстрее, чем у не полностью дифференцированной паренхимы.Этот
вызывает растяжение паренхиматозных клеток, в результате чего межклеточное
образуются пробелы. Они снова частично заполняются при дальнейшем
антиклинальный отдел палисадной паренхимы.
Этот тип меристематической активности связан с
пластинчатая меристема ,
предшественник мезофилла. Дифференциация на типичный
столбчатой паренхимы и губчатой паренхимы происходит за счет
неравномерная скорость роста различных слоев клеток в развивающихся
лист.Активность клеточного деления и растяжения клеток происходит в
разные слои в разное время. Кроме того, клетки
расширяться в разные стороны. Пример: клетки
столбчатая паренхима развивается путем антиклинального деления (под прямым углом
к поверхности листа), таковые сосудистой ткани параллельно. В виде
до тех пор, пока деление и удлинение двух тканей согласованы,
соотношение ячеек остается прежним. Но клетки этого
эпидермис начинается довольно рано в онтогенезе, чтобы разделить больше
быстрее, чем у не полностью дифференцированной паренхимы.Этот
вызывает растяжение паренхиматозных клеток, в результате чего межклеточное
образуются пробелы. Они снова частично заполняются при дальнейшем
антиклинальный отдел палисадной паренхимы.
Дифференцировка нижнего эпидермиса и губчатая паренхима скоординирована несколько более вариабельно. В зависимости от вид представляет собой деление клеток эпидермиса или губчатая паренхима остановилась первой. Деление клетки и клетка удлинения временно отделены друг от друга.Разные скорость роста различных частей развивающегося листа зависит от волнообразное движение роста. Эти роста движения были задокументированы в замедленной съемке. Это показывает впечатляюще, как распространяется рост и расширение листа бука в интенсивные волнообразные движения (W. KAUSCH, Universität Bonn, 1986; производство и дистрибьютор: Institut für Wissenschaftlichen фильм, Геттинген).
© Peter v. Sengbusch — [email protected]
Ассимиляция углекислого газа в некоторых органах и тканях воздушных растений на JSTOR
Абстрактный Характеристики фиксации СО2 ряда зрелых (но не стареющих) тканей и органов (внешние слои зеленого стручка и семенная кожура Vicia faba L.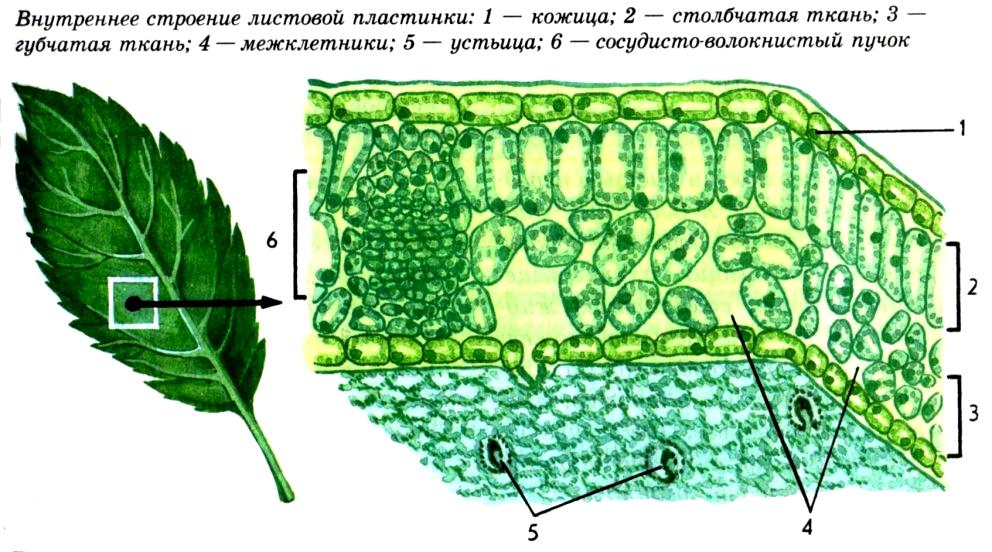 ; наружные слои зеленого стручка и семена Trigonella foenum-graecum L.; внешние слои зеленых плодов Lycopersicon esculentum Mill.) изучали и сравнивали с их соответствующими характеристиками листьев С3. На основе хлорофилла активность фосфоенолпируваткарбоксилазы, яблочного фермента (НАДФ) и малатдегидрогеназы (НАД и НАДФ) была намного выше в нелистовых тканях (за исключением тесты семян V. faba), чем в тканях листа. В целом по белковой основе различия были менее значительными.Все ткани обладали активностью рибулозо-1,5-дифосфаткарбоксилазы, хотя наблюдались большие различия в активности как на белковой, так и на хлорофилловой основе. Соотношение белок: хлорофилл сильно варьировало от ткани к ткани: самое низкое в ткани листа (11,5—14,0) и самое высокое в семенной кожуре V. faba (805,5). Соотношение хлорофилла а:b было в пределах от 2 до 3. Поглощение 14CO2 в темноте ломтиками плодов L. esculentum составляло около ⅓ от поглощения на свету, а основным, первоначально меченым продуктом был малат как на свету, так и в темноте.
; наружные слои зеленого стручка и семена Trigonella foenum-graecum L.; внешние слои зеленых плодов Lycopersicon esculentum Mill.) изучали и сравнивали с их соответствующими характеристиками листьев С3. На основе хлорофилла активность фосфоенолпируваткарбоксилазы, яблочного фермента (НАДФ) и малатдегидрогеназы (НАД и НАДФ) была намного выше в нелистовых тканях (за исключением тесты семян V. faba), чем в тканях листа. В целом по белковой основе различия были менее значительными.Все ткани обладали активностью рибулозо-1,5-дифосфаткарбоксилазы, хотя наблюдались большие различия в активности как на белковой, так и на хлорофилловой основе. Соотношение белок: хлорофилл сильно варьировало от ткани к ткани: самое низкое в ткани листа (11,5—14,0) и самое высокое в семенной кожуре V. faba (805,5). Соотношение хлорофилла а:b было в пределах от 2 до 3. Поглощение 14CO2 в темноте ломтиками плодов L. esculentum составляло около ⅓ от поглощения на свету, а основным, первоначально меченым продуктом был малат как на свету, так и в темноте.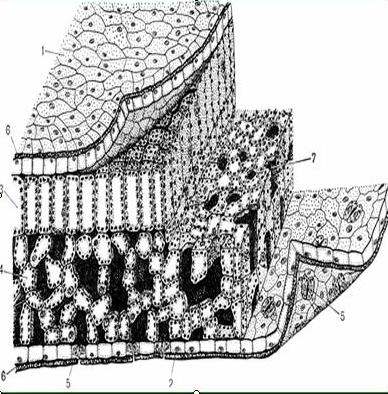 Ни типичный C4-фотосинтез, ни метаболизм крассуловой кислоты не проявлялись в нелистовых тканях, и было сочтено, что присутствовали повышенные уровни активности определенных ферментов для рефиксации и повторного использования вдыхаемого CO2.
Ни типичный C4-фотосинтез, ни метаболизм крассуловой кислоты не проявлялись в нелистовых тканях, и было сочтено, что присутствовали повышенные уровни активности определенных ферментов для рефиксации и повторного использования вдыхаемого CO2.
Planta публикует своевременные и содержательные статьи по всем аспектам биологии растений и представляет оригинальные исследовательские работы по любым видам растений. Области интересов включают биохимию, биоэнергетику, биотехнологию, клеточную биологию, развитие, экологическую и экологическую физиологию, рост, метаболизм, морфогенез, молекулярную биологию, физиологию, взаимодействие растений и микробов, структурную биологию и системную биологию.
Информация об издателе
Springer — одно из ведущих международных научных издательств, выпускающее более 1200 журналов и более
3000 новых книг ежегодно, охватывающих широкий круг предметов, включая биомедицину и науки о жизни, клиническую медицину,
физика, инженерия, математика, информатика и экономика.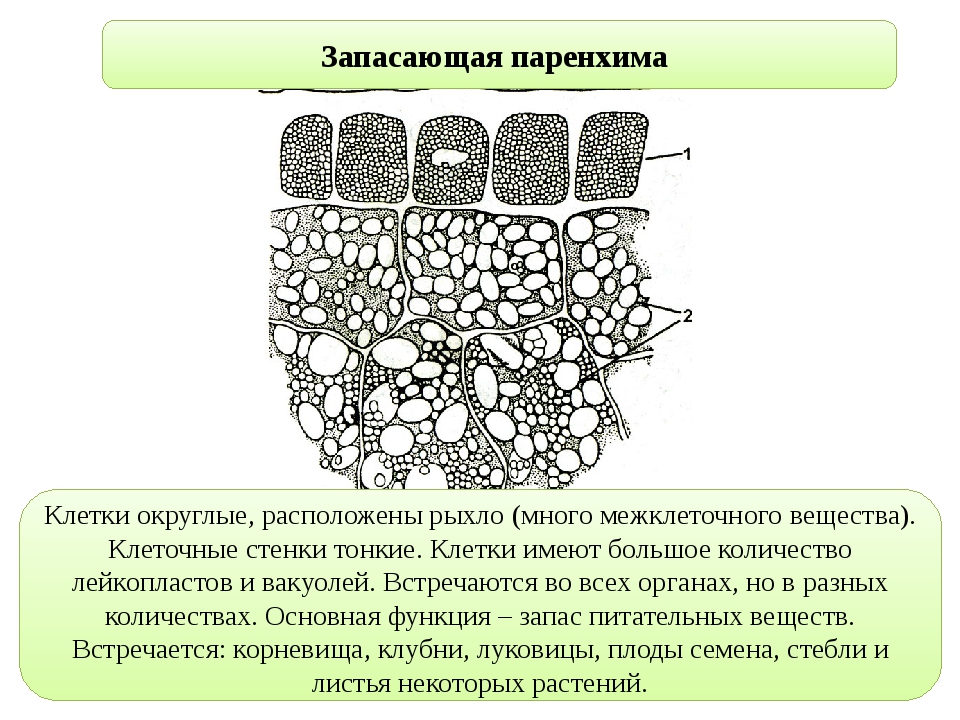
Воздействие повышения концентрации углекислого газа в атмосфере на растения
Эйнсворт, Э.A. Производство риса в условиях меняющегося климата: метаанализ ответов на повышенный уровень углекислого газа и повышенная концентрация озона. Биология глобальных изменений 14 , 1642-1650 (2008).
Эйнсворт, Э. А. и Лонг, С. П. Чему мы научились за 15 лет работы на открытом воздухе CO 2 обогащение (ЛИЦО)? Метааналитический обзор реакций фотосинтеза, свойства навеса и урожайность растений к повышению CO 2 . Новый фитолог 165 , 351-372 (2005).
Эйнсворт, Э. А. и Роджерс А. Реакция фотосинтеза и устьичной проводимости к восхождению (CO 2 ): механизмы и взаимодействие с окружающей средой. Plant, Cell and Environment 30 , 258-270 (2007).
Блум,
A.J., Burger, M. et al. Углерод
диоксид ингибирует ассимиляцию нитратов у пшеницы и арабидопсиса . Наука 328 , 899-903 (2010).
Наука 328 , 899-903 (2010).
Котруфо, М. Ф., Инесон П. и др. Повышенный уровень CO 2 снижает концентрацию азота в тканях растений. Биология глобальных изменений 4 , 43–54 (1998).
Фэн, З., Кобаяши К. и др. Влияние повышенной концентрации озона на рост, физиологию урожай пшеницы ( Triticum aestivum L.): метаанализ. Биология глобальных изменений 14 , 2696-2708 (2008).
де Грааф, Массачусетс, Ван Груниген, К. Дж. и др. др. Взаимодействие между ростом растений и круговоротом питательных веществ в почве при повышенный уровень CO 2 : метаанализ. Глобальный Биология изменений 12 , 2077-2091 (2006).
МГЭИК. Изменение климата 2007: Физическая наука
Основа. Вклад Рабочей группы I в Четвертый доклад об оценке
Межправительственная группа экспертов по изменению климата . Кембридж,
Великобритания: Издательство Кембриджского университета, 2007.
Яблонски, Л. М., Ван, X. и др. Завод репродукция в условиях повышенного содержания CO 2 : метаанализ отчеты по 79 культурным и дикорастущим видам. Новый Фитолог 156 , 9-26 (2002).
Keeling, R. F., Piper, S. C. et al. записей об атмосферном CO2 из точек в сети отбора проб воздуха SIO. В трендах : Сборник данных о глобальных изменениях (Оук-Ридж, ТН: углерод Информация о диоксиде Аналитический центр, Окриджская национальная лаборатория, США Министерство энергетики, 2009 г.).
Лики, A.D.B., Ainsworth, E.A. et al. Надземный CO 2 влияние на отношения углерода, азота и воды в растениях; шесть важные уроки FACE. Журнал Экспериментальная ботаника 60 , 2859-2876 (2009).
Лоладзе,
I. Повышение содержания CO 2 в атмосфере и питание человека: на пути к глобальному
несбалансированная растительная стехиометрия? Тенденции в
Экология и эволюция 17 ,
457-461 (2002).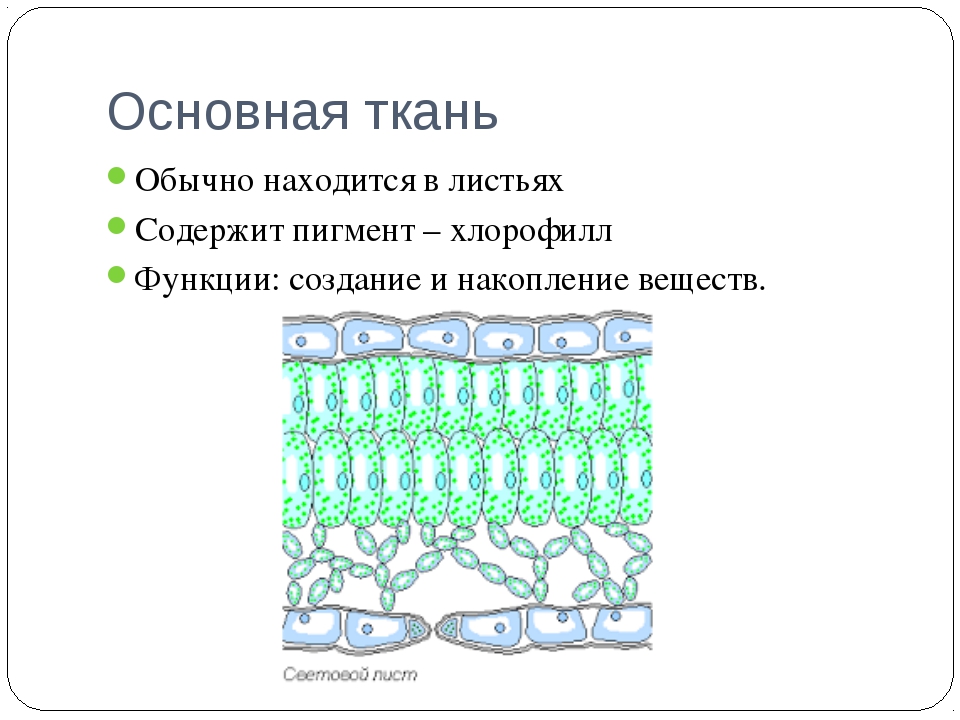
Лонг, С.P., Ainsworth, E.A. и др. Еда для мысль: стимулирование урожайности ниже ожидаемой за счет повышения уровня CO 2 концентрации. Наука 312 , 1918-1921 (2006).
Маршнер, H. Минеральное питание высших растений , 2-е изд. Лондон, СОЕДИНЕННОЕ КОРОЛЕВСТВО: Академическая пресса, 1995.
.Морган, П.Б., Эйнсворт, Э.А. и др. Как работает повышенное воздействие озона на сою? Метаанализ фотосинтеза, роста и урожай. Растения, клетки и окружающая среда 26 , 1317-1328 (2003).
Путер, H. and Navas, M.L. Рост растений и конкуренция при повышенном уровне CO 2 : on победители, проигравшие и функциональные группы. Новый Фитолог 157 , 175-198 (2003).
Роджерс, А.,
Эйнсворт, Э. и др. . Будет повышен
концентрации углекислого газа усиливают преимущества фиксации азота в
бобовые? Физиология растений 151 , 1009-1016 (2009).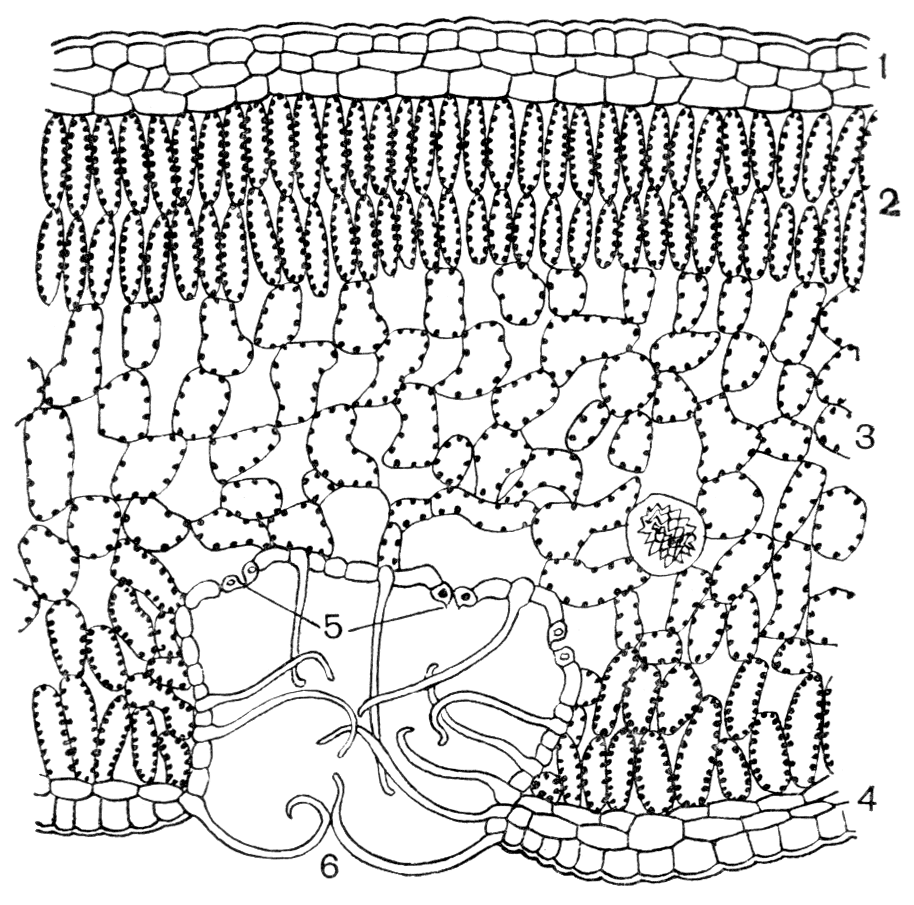
Стайлинг, П. и Корнелиссен, Т. Как влияет повышенный уровень углекислого газа (CO 2 ) влияют на взаимодействие растений и травоядных? Полевой эксперимент и метаанализ CO 2 -опосредованного изменения в химическом составе растений и продуктивности травоядных. Биология глобальных изменений 13 , 1823-1842 (2007).
Тауб, Д., Миллер Б. и др. Эффекты повышенный уровень CO 2 на концентрацию белка в пищевых культурах: a метаанализ. Биология глобальных изменений 14 , 565-575 (2008).
Тауб, Д. Р. и Ван, X. Z. Почему концентрация азота в тканях растений ниже при повышенном уровне CO 2 ? Критическое рассмотрение гипотез. Журнал интегративной биологии растений 50 , 1365-1374 (2008).
Вингарзан Р. Обзор фоновых уровней приземного озона и тенденции. Атмосферная среда 38 , 3431-3442 (2004).
Зиска,
Л.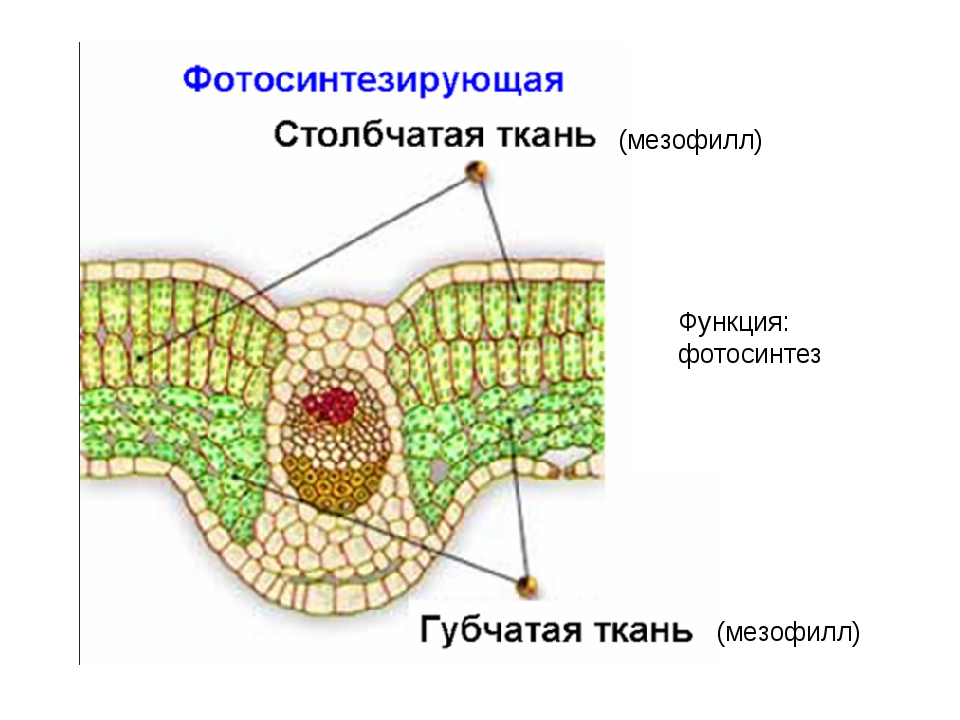 Х. Восходящий атмосферный углекислый газ и биология растений: упущенное из виду.
парадигма. В спорах в науке и
Технология, от климата до хромосом.
изд. Клейнман Д.Л., Клауд-Хансен К.А. и др. (Нью-Рошель: Liebert, Inc., 2008 г.) 379–400.
Х. Восходящий атмосферный углекислый газ и биология растений: упущенное из виду.
парадигма. В спорах в науке и
Технология, от климата до хромосом.
изд. Клейнман Д.Л., Клауд-Хансен К.А. и др. (Нью-Рошель: Liebert, Inc., 2008 г.) 379–400.
Клеточная регуляция ассимиляции железа
Клетки растений, большинства микроорганизмов и животных требуют четко определенных количеств железа для выживания, репликации и дифференцировки. Металл является важным компонентом таких процессов, как синтез ДНК, РНК и хлорофилла; электронный транспорт; кислородный обмен; и фиксации азота. Из-за нерастворимости железа в аэробных средах при нейтральных и щелочных значениях pH клеткам пришлось использовать определенные стратегии для усвоения металла.К ним относятся (1) разработка систем для восстановления ионов трехвалентного железа до более растворимых ионов двухвалентного железа на поверхности клетки, (2) использование небольших молекул-носителей (называемых сидерофорами), которые имеют высокое сродство к ионам трехвалентного железа и рецепторным белкам для ферратированных молекул, и (3) использование трансферрина и других белков, которые могут транспортировать ионы трехвалентного железа.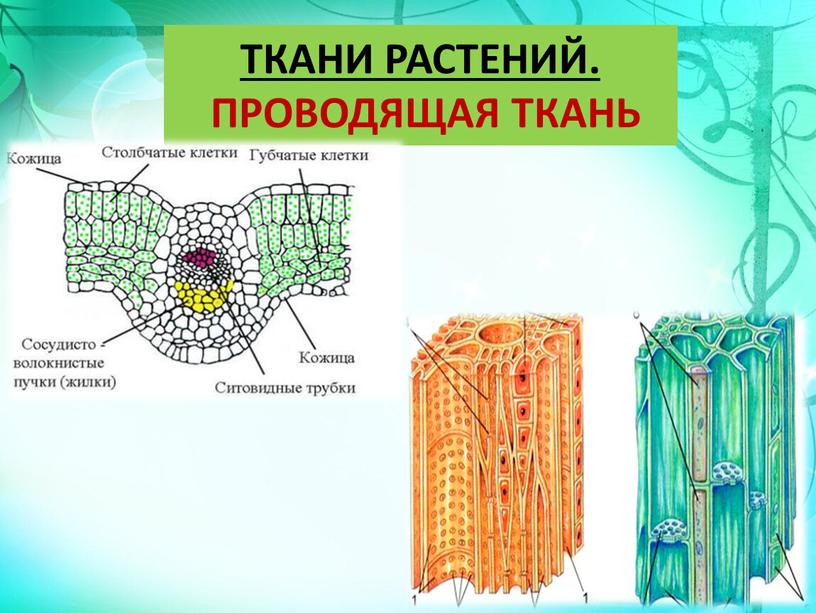 Однако избыточное количество железа токсично, а его внутриклеточная емкость ограничена, а механизмы оттока, как правило, отсутствуют. Таким образом, клетки должны были разработать методы предотвращения чрезмерного накопления металла.К ним относятся использование (1) кислорода для преобразования ионов двухвалентного железа в ионы трехвалентного, (2) небольших молекул, которые могут связывать ионы двухвалентного железа, называемых сидерофраксами, и (3) белков, которые в сочетании с ионами двухвалентного железа подавляют экспрессию генов транспорта железа. Часто один организм может предотвратить рост соседей, ограничивая их доступ к железу. В других случаях клетки помогают друг другу, используя общие системы получения железа или ограничивая приток избыточного железа. Гомеостатический контроль других необходимых микроэлементов также необходим для оптимального функционирования клеток.Тем не менее, поскольку до сих пор больше всего внимания уделялось железу, оно служит моделью минерального обмена.
Однако избыточное количество железа токсично, а его внутриклеточная емкость ограничена, а механизмы оттока, как правило, отсутствуют. Таким образом, клетки должны были разработать методы предотвращения чрезмерного накопления металла.К ним относятся использование (1) кислорода для преобразования ионов двухвалентного железа в ионы трехвалентного, (2) небольших молекул, которые могут связывать ионы двухвалентного железа, называемых сидерофраксами, и (3) белков, которые в сочетании с ионами двухвалентного железа подавляют экспрессию генов транспорта железа. Часто один организм может предотвратить рост соседей, ограничивая их доступ к железу. В других случаях клетки помогают друг другу, используя общие системы получения железа или ограничивая приток избыточного железа. Гомеостатический контроль других необходимых микроэлементов также необходим для оптимального функционирования клеток.Тем не менее, поскольку до сих пор больше всего внимания уделялось железу, оно служит моделью минерального обмена.
