
«Сходства зародышей человека и других позвоночных как доказательство их эволюционного родства»
Конспект урока по теме: «Сходства зародышей человека и других позвоночных как доказательство их эволюционного родства»
Дидактическая цель: Создать условия для формирования знаний о биогенетическом законе как одном из доказательств эволюционного родства позвоночных животных.
Образовательная:
Выявить признаки сходства и различий зародышей разных групп позвоночных животных.
Доказать эволюционное родство между разными группами позвоночных животных.
Развивающая:
Создать условия для развития воображения, речи, внимания, логического мышления;
Анализировать и систематизировать информацию, делать выводы.
Формировать научное мировоззрение.
Совершенствовать навыки самостоятельной работы.
Воспитательная:
Технология: Проблемное обучение, ИКТ.
Тип урока: Комбинированный.
Методы: Изложения и восприятия учебной информации, частично – поисковый, наглядный.
Формы организации познавательной деятельности: Индивидуальная работа, работа в парах, опережающее обучение.
Оборудование: презентация, инструктивные карточки, информационные листы.
Ход урока
Действия учителя
Действия учащихся
Организационный момент
Актуализация опорных знаний:
Что называют онтогенезом?
Из каких периодов складывается индивидуальное развитие организма?
Какие этапы включает в себя эмбриональный этап развития?
Для каких организмов характерен такой тип развития?
Мотивация к изучению новой темы
Ребята, я предлагаю вашему вниманию изображение, глядя на которое вы должны попытаться сформулировать тему урока.

Запишите тему урока: «Сходства зародышей человека и других позвоночных как доказательство их эволюционного родства»
Цель урока: Систематизировать и дополнить знания о сходстве зародышей разных групп позвоночных животных.
Знать: 1. Признаки сходства и различий у позвоночных животных. 2. Биогенетический закон
Уметь: 1. Находить сходство и различия признаков у позвоночных животных.
2. Доказывать эволюционное родство между позвоночными животными и человеком
Учащиеся отвечают на поставленные вопросы.
Учащиеся предлагают темы урока.
Учащиеся записывают в тетрадях тему урока.
Слушают цели и задачи урока.
Учащиеся показывают свои знания по данной теме.
Изучение нового материала:
Как называется наука, которая изучает развитие зародышей?
(Эмбриология – наука о зародышевом развитии).
Из чего начинается развитие организма? ( из оплодотворённой яйцеклетки).
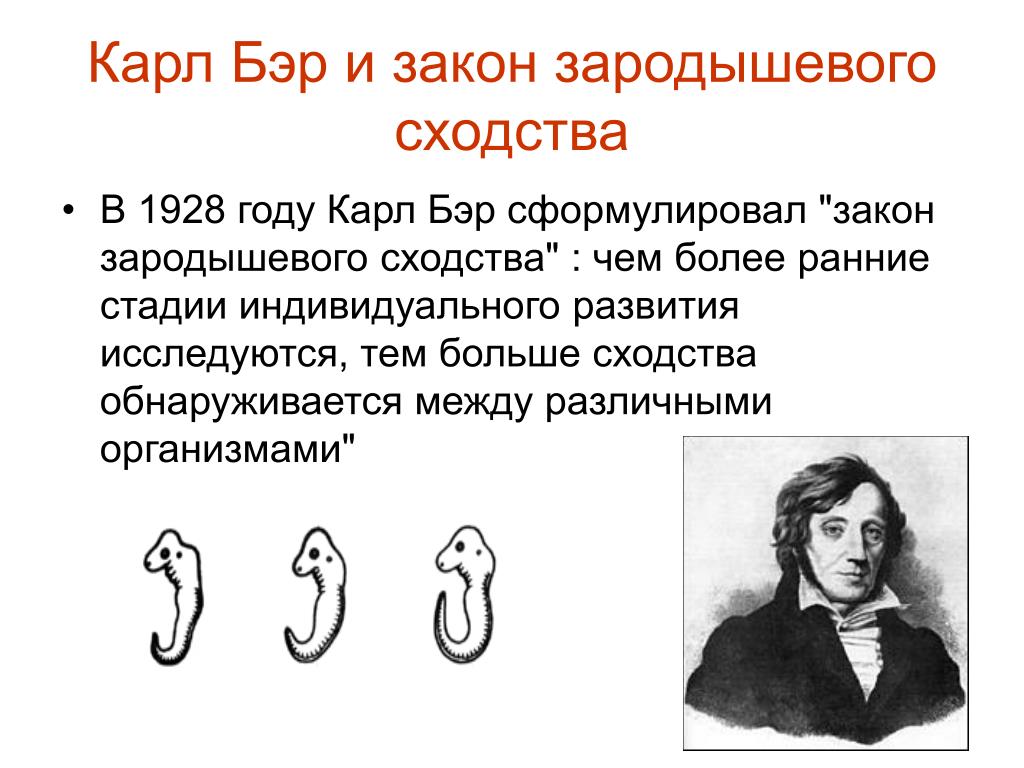
Исследования, проведённые эмбриологами, показали, что зародыши позвоночных животных относящихся к разным группам схожи.
Главные вопросы урока:
В чём проявляется схожесть зародышей?
О чём свидетельствует сходство зародышей?
Какие биологические законы были открыты на основании этого сходства?
Для того, чтобы ответить на эти вопросы, вы сейчас должны выполнить практическую работу.
У вас на столах лежат инструктивные карты, давайте разберем, из каких этапов состоит работа.
Выполнение практической работы.
Обсуждение результатов практической работы. Работа по вопросам:
1.В чём проявляется схожесть зародышей?
2.О чём свидетельствует сходство зародышей?
3.Какие различия наблюдаются у разных групп позвоночных?
На основании этих исследований были сформулированы биологические законы.
Заслушивание сообщения учащегося.
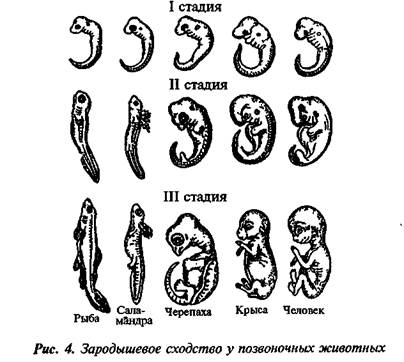
I закон — «Закон зародышевого сходства» В 1828 г. Карл фон Бэр сформулировал закономерность, которую называют Законом Бэра: «Чем более ранние стадии индивидуального развития сравниваются, тем больше сходства удается обнаружить».
II закон – «Биогенетический закон». (Закон Геккеля-Мюллера)
каждое живое существо в своем индивидуальном развитии (онтогенез) повторяет в известной степени формы, пройденные его предками или его видом.
Ответы учащихся.
Учащиеся слушают учителя
Работают с инструктивными картами.
Выполняют практическую работу в тетрадях.
Учащиеся отвечают на вопросы и озвучивают результаты практической работы.
Учащийся выступает с сообщением.
Закрепление
Решение теста с одним правильным ответом.
1.Биогенетический закон был сформулирован:
А – Ч.Дарвиным; Б – К.Бэром; В – Шлейденом и Шваном;
Г – Мюллером и Геккелем.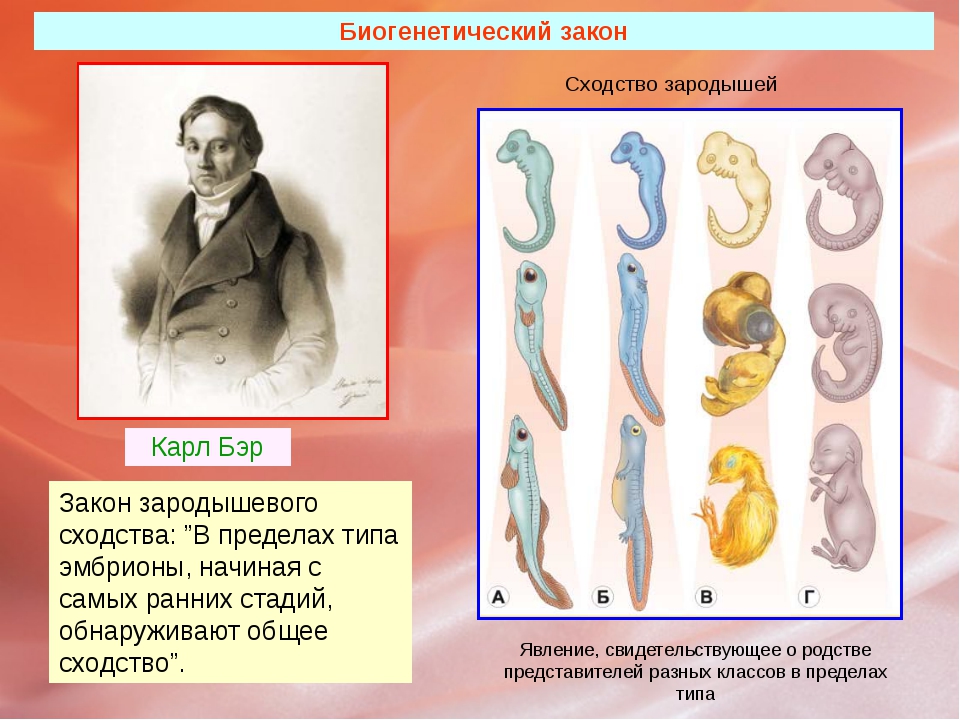
2.Решающим эмбриологическим доказательством эволюции является:
А – сходство деления всех клеток организма;
Б – Сходство в строении скелетов всех позвоночных
В – Сходство ранних стадий эмбрионального развития
Г – Общность строения кровеносной системы позвоночных.
3.К какому закону относится формулировка «Чем более ранние стадии индивидуального развития сравниваются, тем больше сходства удается обнаружить»:
А – Биогенетический закон; Б — Закон Харди — Вайнберга;
В – Закон зародышевого сходства.
4.Какие из признаков схожи между разными группами позвоночных животных:
А- двухкамерное сердце, жабры, хорда; Б – пищеварительная система, конечности, четырёхкамерное сердце. В – головной мозг, выделительная система.
5.О каком эволюционном доказательстве свидетельствует сходство зародышей разных групп позвоночных животных :
А – о палеонтологическом; Б – биогеографическом; В – эмбриональном.
Учащиеся в тетради самостоятельно решают тестовое задание.
Домашнее задание.
Подготовить презентации Причины нарушений в развитии организма»
Алкоголь
Наркотики
Табачная зависимость.
Экология
Лекарственные препараты.
Биогенетический закон
Биология — Биогенетический закон
09 февраля 2011ищу спонсора в москве.
Оглавление:
1. Биогенетический закон
2. Примеры выполнения биогенетического закона
3. Факты, противоречащие биогенетическому закону
4. Связь биогенетического закона с дарвинизмом
5. Научная критика биогенетического закона и дальнейшее развитие учения о связи онтогенеза и филогенеза
6. Критика со стороны креационистов
Биогенетический закон Геккеля-Мюллера: каждое живое существо в своем индивидуальном развитии повторяет в известной степени формы, пройденного его предками или его видом .
Зародыши по Геккелю. Рисунок из книги Ремане, воспроизводящий исходную иллюстрацию Геккеля
Сыграл важную роль в истории развития науки, однако в настоящее время в своем исходном виде не признается современной биологической наукой. По современной трактовке биогенетического закона, предложенной русским биологом А.Н. Северцовым в начале 20 века, в онтогенезе происходит повторение признаков не взрослых особей предков, а их зародышей.
История создания
Фактически «биогенетический закон» был создан ещё задолго до возникновения дарвинизма.
Немецкий анатом и эмбриолог Мартин Ратке в 1825 г. описал жаберные щели и дуги у эмбрионов млекопитающих и птиц один из наиболее ярких примеров рекапитуляции.
В 1824—1826 годах Этьен Серра сформулировал «закон параллелизма Меккеля-Серра»: каждый организм в своем эмбриональном развитии повторяет взрослые формы более примитивных животных.
В 1828 году Карл Максимович Бэр, основываясь на данных Ратке и на результатах собственных исследований развития позвоночных, сформулировал закон зародышевого сходства: «Эмбрионы последовательно переходят в своем развитии от общих признаков типа ко все более специальным признакам.
«Биогенетический закон» как следствие эволюционного развития организмов впервые был сформулирован английским естествоиспытателем Чарльзом Дарвином в его книге «Происхождение видов» в 1859 г: «Интерес эмбриологии значительно повысится, если мы будем видеть в зародыше более или менее затененный образ общего прародителя, во взрослом или личиностном его состоянии, всех членов одного и того же большого класса»
За 2 года до формулировки Эрнстом Геккелем биогенетического закона сходную формулировку предложил на основе своих исследований развития ракообразных работавший в Бразилии немецкий зоолог Фриц Мюллер .
Краткая афористичная формулировка этого закона была дана немецким естествоиспытателем Эрнстом Геккелем в 1866 г. Краткая формулировка закона звучит следующим образом: Онтогенез есть рекапитуляция филогенеза.
Просмотров: 28192
Биогенетический закон Геккеля-Мюллера
Биогенетический закон Геккеля-Мюллера описывает наблюдаемое в живой природе соотношение — онтогенез, то есть персональное развитие каждого живого организма, в определенной степени оно повторяет филогенез — историческое развитие всей группы особей, к которой он относится. Закон сформулировали, как понятно из названия, Э. Геккель и Ф. Мюллер в 60-е годы XIX века независимо друг от друга, и установить первооткрывателя теории сейчас почти невозможно.
Очевидно, что биогенетический закон не был сформулирован сразу. Работе Мюллера и Геккеля предшествовало создание теоретической базы для закона в виде уже обнаруженных явлений и других установленных закономерностей природы. В 1828 году К. Бэр сформулировал так называемый закон зародышевого сходства. Суть его заключается в том, что эмбрионы особей, относящихся к одному биологическому типу, имеют множество схожих элементов анатомического строения. У человека, например, на определенной стадии развития у зародыша имеются жаберные щели и хвост. Характерные отличительные черты в морфологии видов возникают только в ходе дальнейшего онтогенеза. Закон зародышевого сходства во многом определил биогенетический закон: раз эмбрионы различных организмов повторяют стадии развития других особей, они повторяют стадии развития всего типа вообще.
В 1828 году К. Бэр сформулировал так называемый закон зародышевого сходства. Суть его заключается в том, что эмбрионы особей, относящихся к одному биологическому типу, имеют множество схожих элементов анатомического строения. У человека, например, на определенной стадии развития у зародыша имеются жаберные щели и хвост. Характерные отличительные черты в морфологии видов возникают только в ходе дальнейшего онтогенеза. Закон зародышевого сходства во многом определил биогенетический закон: раз эмбрионы различных организмов повторяют стадии развития других особей, они повторяют стадии развития всего типа вообще.
А.Н. Северцовым позднее были внесены определенные поправки к закону Геккеля-Мюллера. Ученый отметил, что во время эмбриогенеза, то есть стадии зародышевого развития, наблюдается сходство между органами именно эмбрионов, а не взрослых особей. Так, жаберные щели у человеческого зародыша схожи с жаберными щелями эмбрионов рыб, но никак не со сформировавшимися жабрами взрослых рыб.
Важно отметить, что одним из наиболее значимых доказательств дарвиновской теории эволюции считается непосредственно биогенетический закон. Формулировка его сама по себе намекает на собственную логическую связь с учением Дарвина. Эмбрион в ходе своего развития проходит через множество различных стадий, каждая из которых напоминает определенные ступени развития природы, отмечаемые с эволюционной точки зрения. Таким образом, каждая все более сложно организованная особь отражает в своем онтогенезе развитие всей живой природы с точки зрения эволюции.
В психологии также существует свой биогенетический закон, сформулированный независимо от биологического. По сути, в психологии выделяется не оформленный закон, а высказанная И. Гербартом и Т. Циллером идея о схожести развития психики ребенка с таковой у человечества вообще. Различные ученые пытались обосновать эту теорию с разных точек зрения. Г. Холл, например, прибегал непосредственно к закону Геккеля-Мюллера. Он говорил о том, что развитие ребенка, в том числе в психологическом плане, задается исключительно биологическими предпосылками и повторяет эволюционное развитие вообще. Так или иначе, на сегодняшний день идея не является однозначно доказанной. В психологии до сих пор не существует биогенетического закона как такового.
Так или иначе, на сегодняшний день идея не является однозначно доказанной. В психологии до сих пор не существует биогенетического закона как такового.
Эмбриологические и сравнительно-морфологические методы
Вопрос. Рассмотрите рис. 50–59. О чём свидетельствуют сходство зародышевого развития позвоночных животных, наличие у организмов гомологичных и аналогичных органов, рудиментов и атавизмов?
Ответ. У всех позвоночных животных наблюдается значительное сходство зародышей на ранних стадиях развития: форма тела, зачатки жабр, хвост, один круг кровообращения и т.д. (закон зародышевого сходства К. Бэра). Однако по мере развития сходство между зародышами различных систематических групп постепенно стирается, и начинают преобладать черты, свойственные их классам, семействам, родам, и, наконец, видам. Биогенетический закон Геккеля-Мюллера гласит: онтогенез есть краткое и быстрое повторение филогенеза, то есть зародыш в индивидуальном развитии повторяет историю развития своего вида.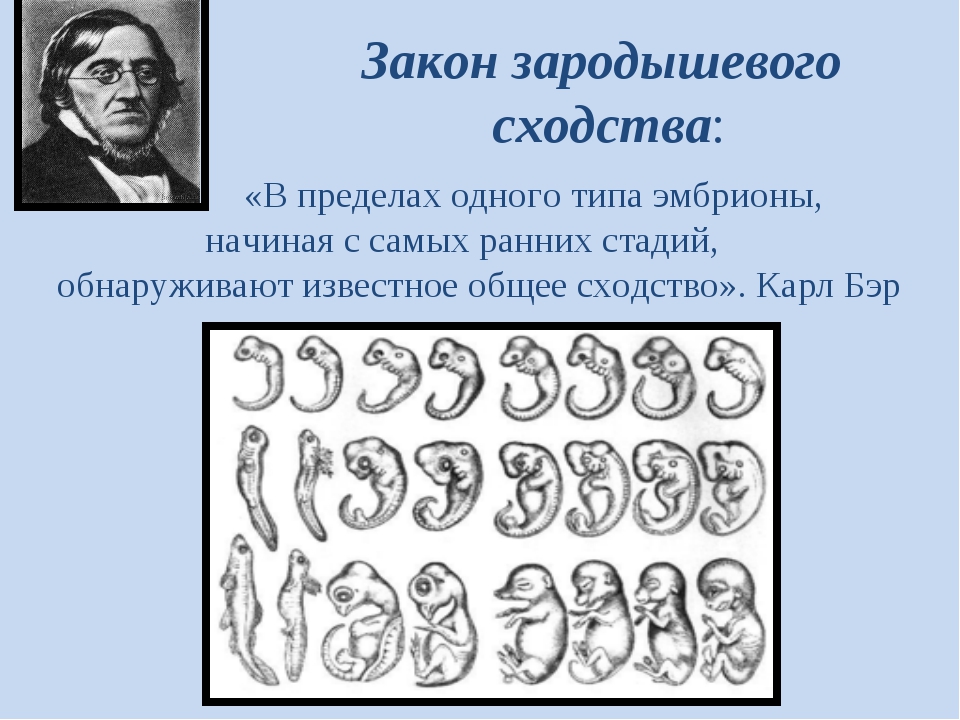
Таким образом, все хордовые животные произошли от единых предков. Об этом же говорит наличие у организмов гомологичных и аналогичных органов, рудиментов и атавизмов
Вопрос 1. В чём сущность биогенетического закона? Кто его авторы? Кем и как была уточнена его первоначальная формулировка? Каково значение закона для изучения эволюции? Приведите соответствующие примеры.
Ответ. В 1864 г. немецкий учёный Фриц Мюллер (1821–1897), проведя ряд наблюдений за развитием ракообразных, предположил, что их предки могли быть похожи на современные формы личинок. Эти исследования позволили ему установить связь между историческим и индивидуальным развитием организмов и сделать важный вывод: индивидуальное развитие (онтогенез) повторяет историческое развитие вида (филогенез). Позднее Эрнст Геккель (1834–1919) сформулировал это обобщение в виде биогенетического закона: онтогенез особи есть краткое и быстрое повторение филогенеза. Закон Геккеля – Мюллера имеет большое значение для установления родственных связей между организмами.
Позднее формулировка биогенетического закона была дополнена и уточнена русским учёным Алексеем Николаевичем Северцовым (1866–1936). Он установил, что в онтогенезе повторяются не взрослые формы предков, а их эмбриональные стадии, причём в процессе индивидуального развития некоторых организмов отдельные стадии могут выпадать. Кроме того, возникают новообразования, которые отсутствуют у зародышей предков. Таким образом, в индивидуальном развитии одних организмов закладываются новые эволюционные пути развития других. Биогенетический закон позволяет использовать данные, полученные при изучении развития зародышей, для понимания хода эволюции. Но поскольку короткий онтогенез не может воспроизвести все стадии филогенеза, длившегося сотни миллионов лет, то в нем воспроизводятся лишь отдельные эволюционные этапы. У позвоночных на ранних стадиях эмбрионального развития наблюдается большое сходство между представителями различных классов: у всех имеется головной туловищный и хвостовой отделы, зачатки конечностей и жабр. Но в дальнейшем у рыб из зачатков жаберных дуг образуется жаберный аппарат, а у млекопитающих – совершенно иные органы (хрящи гортани и трахеи). Таким образом, в онтогенезе млекопитающих повторяется не строение взрослых рыб, а строение закладок эмбриональных органов.
Но в дальнейшем у рыб из зачатков жаберных дуг образуется жаберный аппарат, а у млекопитающих – совершенно иные органы (хрящи гортани и трахеи). Таким образом, в онтогенезе млекопитающих повторяется не строение взрослых рыб, а строение закладок эмбриональных органов.
Вопрос 2. Используя рис. 50, перечислите черты внешнего сходства и различия у зародышей позвоночных животных на разных стадиях развития.
Ответ. Эмбрионы различных классов и видов позвоночных животных обнаруживают в пределах типа большое сходство. Наибольшее сходство имеют зародыши на ранних стадиях развития: общие признаки типа формируются в ходе эмбриогенеза раньше, чем специальные. Так, все эмбрионы позвоночных на первой стадии имеют жаберные щели и двухкамерное сердце. Затем появляются черты строения, характерные для каждого класса, и лишь на более поздних стадиях формируются особенности конкретного вида
Вопрос 3. У высших позвоночных животных в эмбриогенезе закладываются жаберные щели с жаберными артериями и венами, которые затем зарастают и у взрослого организма отсутствуют.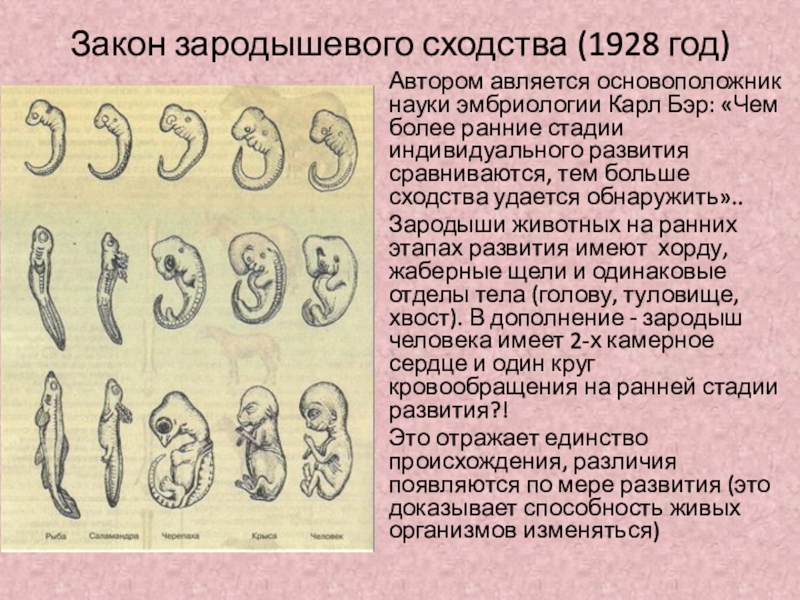 Естественно, что для животных, дышащих лёгкими, жаберные щели не могут иметь приспособительного значения. Объясните, почему из поколения в поколение у эмбрионов сохраняется этот признак.
Естественно, что для животных, дышащих лёгкими, жаберные щели не могут иметь приспособительного значения. Объясните, почему из поколения в поколение у эмбрионов сохраняется этот признак.
Ответ. Повторение структур, характерных для предков, в эмбриогенезе потомков названо рекапитуляциями. Рекапитулируют не только морфологические признаки – хорда, закладки жаберных щелей и жаберных дуг- у всех хордовых, но и особенности биохимической организации и физиологии.
У высших позвоночных рекапитуляции артериальных жаберных дуг происходят с особенностями: все шесть пар дуг никогда не существуют одновременно. В то время, когда две первые дуги закладываются, а затем перестраиваются, последние пары сосудов еще не начинают формироваться. Кроме того, пятая артериальная дуга уже закладывается в виде рудиментарного сосуда, присоединенного обычно к 4-й паре, и редуцируется очень быстро. Рекапитуляции проявляются и в эмбриональном развитии крупных вен.
Генетическая основа рекапитуляции заключена в единстве механизмов генетического контроля развития, сохраняющемся на базе общих генов регуляции онтогенеза, которые достаются родственным группам организмов от общих предков.
Вопрос 4. Что такое гомологичные и аналогичные органы? В чём их сходство и различие? Перечертите в тетрадь и заполните таблицу.
Ответ. Гомологичные органы (от греч. homologia – соответствие, согласие) имеют сходный план строения и развиваются из одинаковых зародышевых зачатков. Они выполняют как сходные, так и различные функции. Примером гомологичных органов являются передние конечности представителей разных групп позвоночных животных, имеющие общий план строения независимо от выполняемых функций Аналогичные органы (от греч. analogia – соответствие, сходство, подобие) – морфологически сходные органы, выполняющие одинаковые функции, но не имеющие единого плана строения и развивающиеся из разных зародышевых зачатков. Они свидетельствуют о сходстве приспособлений к одинаковым условиям среды, возникающих у разных организмов в ходе эволюции. Примеры аналогичных органов у животных – жабры рака и рыбы, крылья бабочки и птицы, бегательные конечности собаки и жука.
Гомология и аналогия у животных и растений
Вопрос 5. Что такое рудименты и атавизмы? Приведите примеры рудиментов и атавизмов. В чём их принципиальное различие?
Ответ. Рудиментарные органы, или рудименты (от лат. rudimentum – зачаток, первооснова) – это сравнительно упрощённые или недоразвитые образования, утратившие своё первоначальное значение. Они закладываются в период эмбрионального развития организма, но полностью не развиваются. Рудименты тазового пояса кита и конечностей питона подтверждают факт происхождения этих животных от наземных предков с развитыми конечностями. Однако рудиментарные тазовые косточки кита выполняют совершенно иную функцию – они связаны с мышцами мочеполовой системы. К рудиментам относят также грифельные косточки в конечности лошади, первый и третий пальцы в крыле птицы, волосяной покров на теле человека.
Примером рудиментов у растений могут служить цветочные чешуи и плёнки в цветке злаков, редуцированный околоцветник у ветроопыляемых растений. Рудиментарными листьями являются чешуйки на корневищах ландыша, купены. Атавизм (от лат. atauus – предок) – явление возврата к признакам предков, проявляющееся у отдельных особей. Например, появление трёхпалой конечности у жеребят, многососковость у человека.
Рудиментарными листьями являются чешуйки на корневищах ландыша, купены. Атавизм (от лат. atauus – предок) – явление возврата к признакам предков, проявляющееся у отдельных особей. Например, появление трёхпалой конечности у жеребят, многососковость у человека.
Принципиальное различие заключается в том, что рудименты имеются всегда и у всех организмов, а вот атавизмы проявляются только у отдельных особей.
Вопрос 6. Объясните с позиций молекулярной биологии и генетики наличие у организмов рудиментарных органов и появление атавизмов.
Ответ. Рудименты заложены в генетический аппарат всех ныне существующих организмов и проявляются в онтогенезе. Наличие рудиментов объясняется постепенной утратой нормально развитых у предков органов, которые в процессе эволюции потеряли своё первоначальное значение, недоразвились или видоизменились.
Атавизмы. Причина появления атавизмов кроется в наличии у особей «генов предков», ответственных за данный признак. В норме их действие блокируется генами-репрессорами, но когда репрессоры перестают выполнять свои функции, гены активизируются, что приводит к появлению атавизмов. Иногда это бывает связано с общей задержкой онтогенеза. В отличие от рудиментов, которые встречаются у всех особей определённого вида, атавизмы появляются внезапно только у единичных представителей.
В норме их действие блокируется генами-репрессорами, но когда репрессоры перестают выполнять свои функции, гены активизируются, что приводит к появлению атавизмов. Иногда это бывает связано с общей задержкой онтогенеза. В отличие от рудиментов, которые встречаются у всех особей определённого вида, атавизмы появляются внезапно только у единичных представителей.
(PDF) МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ РОССИЙСКОЙ ФЕДЕРАЦИИ
из паразитических растений (рода Cuscuta) в группу сорных видов-хозяев рода
Plantago. Напротив, растение-паразит раффлезия, как установлено, получило
митохондриальные гены от своего хозяина – лианы. Поскольку известно более 4
тысяч видов растений-паразитов (среди которых есть и многолетние), то
предполагается, что у продуцентов в системах «паразит-хозяин» (в ауто- и
демоценозах) ГПГ играет важную эволюционную роль (Шестаков, 2009).
У многоклеточных животных (в отличие от простейших и растений)
достоверных фактов ГПГ отмечено немного, и они касаются передачи генов от
симбиотических бактерий (в основном кишечных) к хозяевам (например, круглым
червям) (см. Шестаков, 2009).
Шестаков, 2009).
Косвенные генетические связи в аутогеноме проявляются в форме
коэволюционного развития как в геноме хозяев, так и в геномах их симбионтов
особых генных комплексов, участвующих со стороны хозяев в адаптивной
регуляции жизнедеятельности симбионтов, а со стороны последних – в
приспособлении к обитанию в теле хозяев и получении от них необходимых
вещественных и энергетических ресурсов (Проворов, Долгих, 2006; Проворов,
2009). При этом, например, известно, что паразитирующие у членистоногих и
нематод бактерии рода Wolbachia и бактерии-мутуалисты (в частности, Buchnera
aphidicola), развивающиеся в бактериоцитах ряда насекомых, передаются
потомкам трансовариально. Это обусловливает особую форму коэволюции
партнеров по симбиозу, в том числе на генетическом уровне. У млекопитающих
передача симбионтов потомкам происходит путем груминга или поедания
фекалий, а у растений, размножающихся вегетативно – через клоны. В результате
В результате
в таких демоценозах происходит вертикальная передача симбионтов, а с ними – и
генетических элементов, приобретаемых партнерами под влиянием симбиогенеза.
Фенотипы хозяина и симбионтов интегрированы в аутофеном (Савинов,
2009), то есть образуют динамичную, симбиогенную структурно-функциональную
подсистему аутоценоза, обусловливающую все его эмерджентные (новые)
фенотипические признаки: морфологические, анатомические, физиологические,
биохимические и этологические. Причем во многих случаях аутофеномы
формируются не только с участием симбиотических прокариот и Protozoa, но и
Metazoa, которые часто являются экто- и эндосимбионтами беспозвоночных
(губок, кишечнополостных, червей, моллюсков, иглокожих, членистоногих),
асцидий, всех позвоночных, в том числе людей. Аутофеномы, развивающиеся при
этом, обнаруживают биохимическую, физиологическую, анатомо-
морфологическую и этологическую эмерджентность.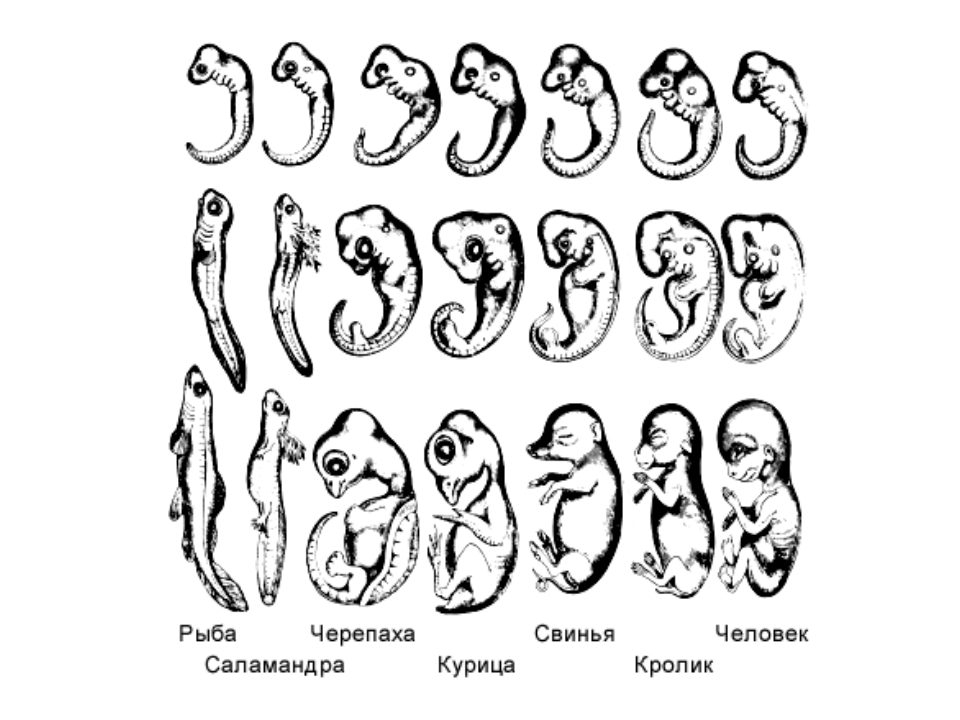 Например, у моллюсков как
Например, у моллюсков как
хозяев трематод, меняется экспрессия генов, обеспечивающих синтез белков,
участвующих в иммунных реакциях, модифицируется кислородный, углеводный,
белковый и липидный обмен, меняется величина среднесуточного рациона и
элективность питания, развивается тахикардия, вакуолизируются гепатоциты,
угнетается репродуктивная способность (уменьшаются количество и размеры яиц,
происходит паразитарная кастрация – атрофия гонад, уменьшение размеров и
Закон фон Бэра | Encyclopedia.com
Ранние стадии развития признака, как правило, более схожи между родственными видами, чем более поздние стадии. Это означает, что большинство характеристик, которые различают таксонов и которые обычно используются для различения видов, представляют собой более поздние модификации принципиально сходного плана развития. Закон фон Бэра гласит, что структуры, формирующиеся на ранних этапах развития, более широко распространены среди групп организмов, чем структуры, возникающие в более поздних стадиях развития.
Этот закон эмбриологии назван в честь немецкого биолога девятнадцатого века Карла Эрнста фон Бэра, который впервые сформулировал его. Хотя фон Бэр не был эволюционистом, его концепция усиления дифференциации между видами в течение онтогенеза , или эмбрионального развития, хорошо согласуется с эволюционным взглядом на эмбриологию. Его закон исходит из первых двух из четырех обобщений, сделанных им в 1828 г.:
- Общие признаки, общие для большой группы животных, появляются у зародыша раньше, чем специализированные.
- В ходе онтогенеза развитие отдельных эмбриональных признаков идет от общего к специализированному.
- Каждый зародыш данного вида, вместо того чтобы пройти через взрослые стадии других животных, по мере развития онтогенеза все более и более удаляется от них (в прямом противоречии с недействительным биогенетическим законом Эрнста Геккеля, обсуждаемым ниже).
- Следовательно, ранний зародыш высшего животного никогда не похож на взрослую особь низшего животного, а только подобен его раннему зародышу.
Закон фон Бэра гласит, что генерализованное состояние присутствует у раннего эмбриона. Это генерализованное состояние уступает место более специализированным состояниям по мере развития зародыша, что можно увидеть при непосредственном наблюдении за зародышами. Другими словами, ранняя стадия куриного эмбриона может быть признана представителем типа позвоночных, но не каким-либо конкретным субтаксоном. Позже, по мере продолжения структурной специализации, его можно распознать как члена класса Aves и, наконец, как представителя определенного вида, Gallus domesticus.
Чарльз Дарвин признал, что закон фон Бэра обеспечивает связь между эмбриологией и биологической эволюцией, а именно, что примитивные признаки имеют тенденцию к обобщению, а производные признаки — к специализации. Дарвин понял, что эволюционные изменения можно вывести из изменений в развитии.
Примером связи между эмбриональным развитием и историей эволюции является развитие конечностей позвоночных. Структуры конечности развиваются в проксимодистальной последовательности или от ближнего к наиболее удаленному от тела: первыми развиваются плечевые кости, а последними — кости пальцев.Двоякодышащие рыбы и латимерии, дальние живущие родственники предковых четвероногих, имеют плавники с хорошо развитыми «плечами», но их придатки не содержат костей , гомологичных пальцам четвероногих (сходство их у разных таксонов вытекает из их общего происхождения в общий предок). У ранних четвероногих пальцы появляются как специализированные, производные черты. Морфология конечностей, или форма, переходит от генерализованной к специализированной по мере того, как мы поднимаемся по филогенетическому , или эволюционному, древовидному и конечностному развитию эмбрионов продвинутых четвероногих параллельно эволюционной истории этих четвероногих.
Структуры конечности развиваются в проксимодистальной последовательности или от ближнего к наиболее удаленному от тела: первыми развиваются плечевые кости, а последними — кости пальцев.Двоякодышащие рыбы и латимерии, дальние живущие родственники предковых четвероногих, имеют плавники с хорошо развитыми «плечами», но их придатки не содержат костей , гомологичных пальцам четвероногих (сходство их у разных таксонов вытекает из их общего происхождения в общий предок). У ранних четвероногих пальцы появляются как специализированные, производные черты. Морфология конечностей, или форма, переходит от генерализованной к специализированной по мере того, как мы поднимаемся по филогенетическому , или эволюционному, древовидному и конечностному развитию эмбрионов продвинутых четвероногих параллельно эволюционной истории этих четвероногих.
Еще одним примером действия закона фон Бэра является разработка нотокорда . Все позвоночных развивают спинной хрящевой стержень, называемый хордой, и жаберные мешочки в области глотки или горла. Оба являются признаками раннего развития и являются общими для многих видов. Однако позже в процессе развития эти признаки утрачиваются и заменяются признаками, характерными для разных групп позвоночных.
Оба являются признаками раннего развития и являются общими для многих видов. Однако позже в процессе развития эти признаки утрачиваются и заменяются признаками, характерными для разных групп позвоночных.
Таким образом, ранние эмбрионы разных видов позвоночных удивительно похожи.Например, первые эмбрионы рыб, саламандр, черепах, цыплят, свиней, коров, кроликов и людей кажутся похожими, однако каждый вид следует своей собственной траектории эмбрионального развития (последовательности событий во время развития), чтобы развить отличительные черты.
Мутации , нарушающие раннее развитие, обычно смертельны, поскольку могут привести к радикальным изменениям в последующем развитии. Из-за этих ограничений развития ранние стадии развития с меньшей вероятностью изменятся с течением времени эволюции.Эти ранние стадии, вероятно, будут сходными среди таксонов, а различия между ними, скорее всего, возникнут позже, в онтогенезе или эмбриональном развитии.
Другой немецкий биолог девятнадцатого века, Эрнст Геккель, предложил то, что он считал законом эволюции.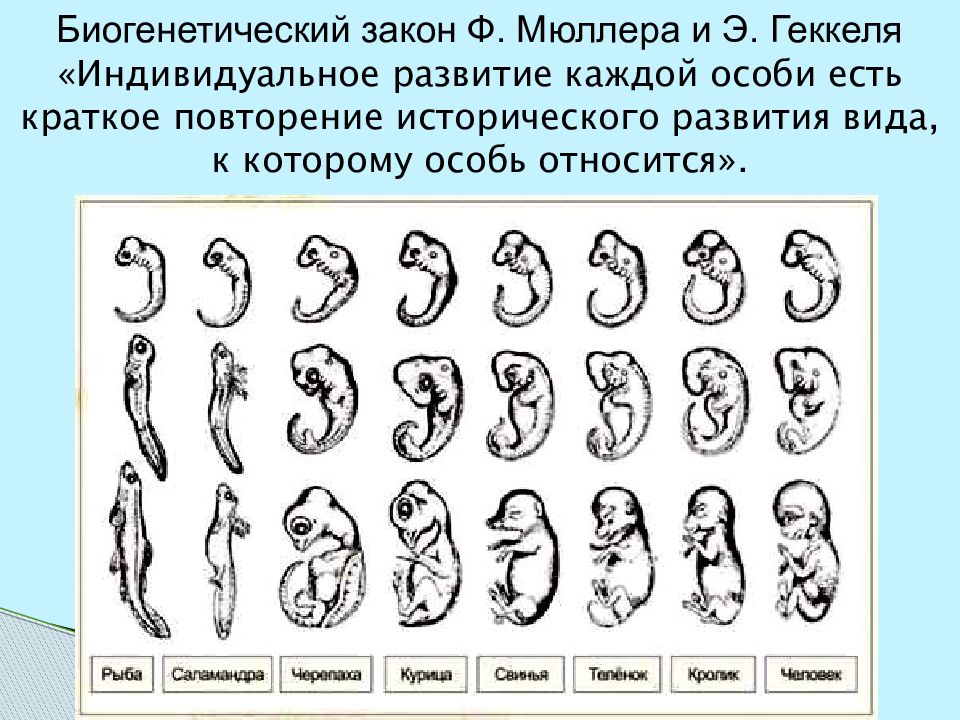 Хотя закон Геккеля недействителен, он широко известен и привел к известной фразе «онтогенез резюмирует филогенез». Таким образом, биогенный закон Геккеля утверждает, что эмбриональное развитие повторяет историю эволюции. Он говорил, что эмбрионы повторяют взрослые стадии предковых видов в хронологическом порядке от примитивных до самых поздних, а изменения добавляются только на последних стадиях, называемых терминальным добавлением.
Хотя закон Геккеля недействителен, он широко известен и привел к известной фразе «онтогенез резюмирует филогенез». Таким образом, биогенный закон Геккеля утверждает, что эмбриональное развитие повторяет историю эволюции. Он говорил, что эмбрионы повторяют взрослые стадии предковых видов в хронологическом порядке от примитивных до самых поздних, а изменения добавляются только на последних стадиях, называемых терминальным добавлением.
В некоторых случаях онтогенез признака действительно проходит стадии, очень похожие на взрослые формы предковых видов. Однако, хотя биогенный закон Геккеля описывает возможную закономерность эволюции, он не дает причинного объяснения этой закономерности. Биогенетический закон в конце концов утратил доверие с появлением экспериментальной эмбриологии и менделевской генетики . Эмбриологические исследования показали, что возможны многие типы изменения времени развития (гетерохрония) и что разные части организма могут различаться по скорости развития. Генетические исследования показали, что 90 003 гена 90 004 могут вызывать изменения на любой стадии развития, так что терминальное добавление не было единственной возможностью.
Генетические исследования показали, что 90 003 гена 90 004 могут вызывать изменения на любой стадии развития, так что терминальное добавление не было единственной возможностью.
см. также Эмбриональное развитие.
Эндрю Г. Глюзенкамп
Библиография
Футуйма, Дуглас Дж. Эволюционная биология. Сандерленд, Массачусетс: Sinauer Associates, Inc., 1986.
Гулд, Стефан Дж. Онтогенез и филогения. Кембридж, Массачусетс: Belknap Press, 1977.
Рафф, Рудольф А. Форма жизни. Чикаго: University of Chicago Press, 1996.
Рафф, Рудольф А. и Томас К. Кауфман. Эмбрионы, гены и эволюция: генетическая основа эволюционных изменений. New York: Macmillan, 1983.
Эрнст Геккель и биогенетический закон (обоснованное мнение)
Гибельный союз эмбриологии и эволюционной биологии был создан во второй половине девятнадцатого века немецким эмбриологом и философом Эрнстом Геккелем.Основываясь на предположении, что законы возникновения видов на этой планете (филогенез) тождественны законам развития особей вида (онтогенез), он рассматривал взрослые организмы как зародышевые стадии более развитых организмов.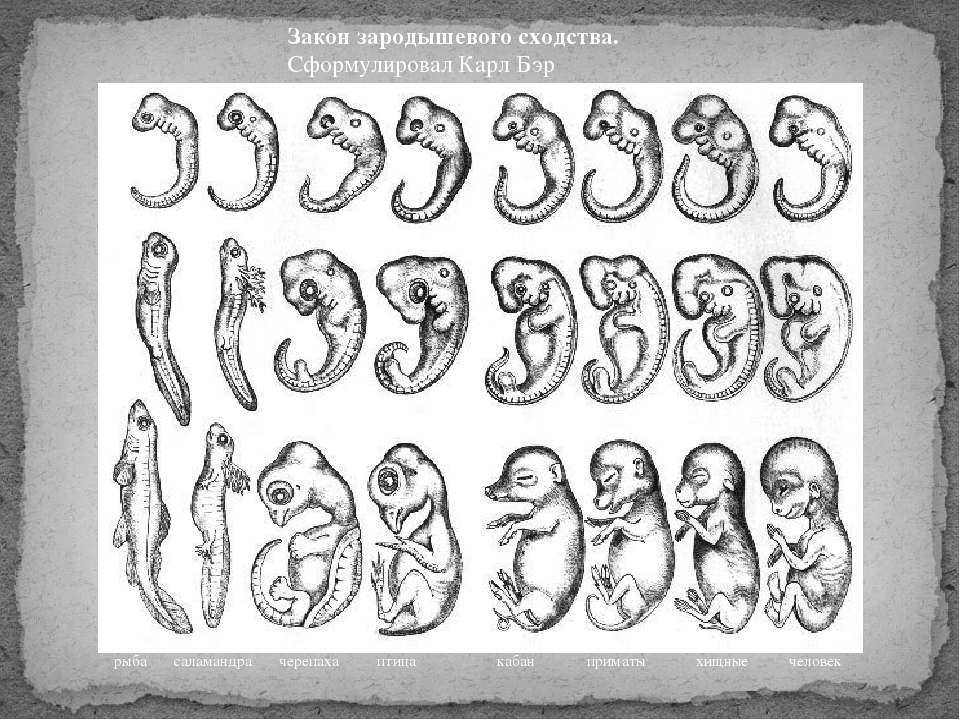 Эта точка зрения была резюмирована в его «Биогенетическом законе» : Онтогенез резюмирует филогенез. Другими словами, развитие продвинутых видов проходило через стадии, представленные взрослыми организмами более примитивных видов. С этой точки зрения создание новых типов — это шаг к завершению человеческого развития.В более ранние эпохи происходили только начальные стадии этого развития, производившие протистов и книдарий. Позже последовательно добавляются новые стадии, пока не разовьется человек. Согласно Геккелю, трех правил было достаточно, чтобы объяснить, как этот развивающийся онтогенез мог породить новые виды. Во-первых, был закон соответствия . Человеческая зигота, например, была представлена «взрослой» стадией протистов; колониальные протисты представляли продвижение развития к стадии бластулы; стадия жаберных щелей эмбрионов человека была представлена взрослыми рыбами.Геккель даже постулировал вымерший организм, Gastraea , двухслойный мешок, соответствующий гаструле, которую он считал предком всех многоклеточных видов (Haeckel 1867, 1879; см.
Эта точка зрения была резюмирована в его «Биогенетическом законе» : Онтогенез резюмирует филогенез. Другими словами, развитие продвинутых видов проходило через стадии, представленные взрослыми организмами более примитивных видов. С этой точки зрения создание новых типов — это шаг к завершению человеческого развития.В более ранние эпохи происходили только начальные стадии этого развития, производившие протистов и книдарий. Позже последовательно добавляются новые стадии, пока не разовьется человек. Согласно Геккелю, трех правил было достаточно, чтобы объяснить, как этот развивающийся онтогенез мог породить новые виды. Во-первых, был закон соответствия . Человеческая зигота, например, была представлена «взрослой» стадией протистов; колониальные протисты представляли продвижение развития к стадии бластулы; стадия жаберных щелей эмбрионов человека была представлена взрослыми рыбами.Геккель даже постулировал вымерший организм, Gastraea , двухслойный мешок, соответствующий гаструле, которую он считал предком всех многоклеточных видов (Haeckel 1867, 1879; см. Gould 1977a).
Gould 1977a).
Во-вторых, существовал закон конечного сложения. Эмбрион развил новый вид, добавив шаг в конце предыдущих. С такой точки зрения, люди развились, когда эмбрион следующей высшей обезьяны добавил новую стадию. Это обеспечило линейную, а не ветвящуюся филогению.Это критически важный отход от того, что мы обычно считаем дарвиновской эволюцией.
Существовал также закон об усечении, , согласно которому предыдущее развитие можно было сократить в ракурсе. Этот закон был необходим для того, чтобы время беременности не было огромным. Это также было необходимо, так как эмбриологи не наблюдали все эти стадии у всех животных.
Это представление об онтогенезе, повторяющем филогенез, не было дарвинизмом. На самом деле синтез Геккеля был попыткой объединить работы Дарвина, Ламарка и Гёте.В дарвинизме современные виды рассматриваются как имеющие общего предка. В результате получается многоветвистый «куст». (Также использовалась метафора дерева, но у деревьев есть центральная ось, на которой ученые часто помещают линию, ведущую к H.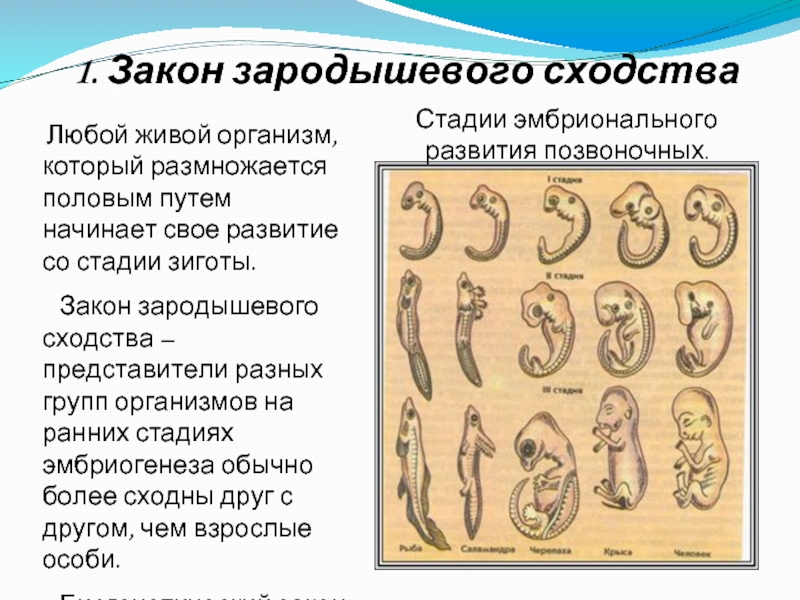 sapiens. ). Люди не «выше» шимпанзе, но имеют предка, от которого оба группы разошлись. В схеме Геккеля животные переходят на новые уровни, добавляя стадии к существующему эмбриональному развитию. Люди были буквально на вершине.Интересно, что фон Бэр (1828) опроверг «биогенетический закон» еще до того, как его изобрел Геккель. Высмеивая доэволюционные формы этого закона, фон Бэр фантазировал о том, что произошло бы, если бы птицы писали учебники по эмбриологии. « На их теле нет настоящего пера, а только тонкие стержни пера», — воображал он, что пишут ученые-птицы. «Мы, как птенцы в гнезде, более продвинуты, чем они когда-либо будут».
sapiens. ). Люди не «выше» шимпанзе, но имеют предка, от которого оба группы разошлись. В схеме Геккеля животные переходят на новые уровни, добавляя стадии к существующему эмбриональному развитию. Люди были буквально на вершине.Интересно, что фон Бэр (1828) опроверг «биогенетический закон» еще до того, как его изобрел Геккель. Высмеивая доэволюционные формы этого закона, фон Бэр фантазировал о том, что произошло бы, если бы птицы писали учебники по эмбриологии. « На их теле нет настоящего пера, а только тонкие стержни пера», — воображал он, что пишут ученые-птицы. «Мы, как птенцы в гнезде, более продвинуты, чем они когда-либо будут».
Наблюдая за развитием, фон Бэр заметил, что эмбрионы никогда не проходят через взрослые стадии других животных.Однако есть стадии, которые родственные эмбрионы разделяют. Все зародыши позвоночных проходят стадию, на которой появляются зародышевые жаберные щели. Рыбы превращают их в настоящие жабры, а у других позвоночных щели становятся частью челюстного или ушного аппарата. Но лягушка или человеческий зародыш никогда не проходят стадию, на которой они приобретают строение взрослой рыбы. Однако, хотя фон Бэр и другие дискредитировали понятие перепросмотра, оно стало одним из самых популярных в биологии. Гулд (1977a, b) показал, что, хотя рекапитуляция имеет ограниченное значение при изучении образования родственных видов, она не является общим явлением.Однако рекапитуляционизм стал одной из центральных парадигм биологии. Когда его в конце концов отвергли, представление о том, что эмбриология была важной движущей силой эволюции, также было отвергнуто. (Действительно, половина книги Стивена Дж. Гулда 1977 года « Онтогенез и филогения » посвящена изгнанию призрака Геккеля, чтобы мы могли обсуждать эволюционную биологию развития, не прибегая к биогенетическим законам.)
Но лягушка или человеческий зародыш никогда не проходят стадию, на которой они приобретают строение взрослой рыбы. Однако, хотя фон Бэр и другие дискредитировали понятие перепросмотра, оно стало одним из самых популярных в биологии. Гулд (1977a, b) показал, что, хотя рекапитуляция имеет ограниченное значение при изучении образования родственных видов, она не является общим явлением.Однако рекапитуляционизм стал одной из центральных парадигм биологии. Когда его в конце концов отвергли, представление о том, что эмбриология была важной движущей силой эволюции, также было отвергнуто. (Действительно, половина книги Стивена Дж. Гулда 1977 года « Онтогенез и филогения » посвящена изгнанию призрака Геккеля, чтобы мы могли обсуждать эволюционную биологию развития, не прибегая к биогенетическим законам.)
Социальное использование синтеза Геккеля
Даже в большей степени, чем в биологии, «биогенетический закон» Геккеля был некритически адаптирован многими вновь формирующимися общественными науками. Ранние антропологи придерживались точки зрения, что другие культуры были «примитивными» в эмбриологическом смысле, поскольку их развитие остановилось на уровне нашего собственного. Действительно, для определения такой культуры до сих пор используется слово «недоразвитая». Поскольку эволюция представляла собой последовательное добавление к вершине дерева, различные расы можно было расположить сверху вниз. (Действительно, их нужно было бы упорядочить линейно, поскольку это не модель с разветвленной цепью.) Ранее несколько историков науки упоминали, что Геккель был антисемитом и что его биология использовалась Третьим рейхом.Эти претензии были повторены на этом веб-сайте. Хотя его биология, безусловно, пыталась классифицировать человеческие группы по расовым характеристикам и использовалась для оправдания притязаний одной этнической группы на превосходство над другими, она не была явно антисемитской. Не был Геккель и антисемитом. Боб Ричардс (2007) обнаружил, что произошла путаница в идентичностях (среди прочего) с другим (и более молодым) Эрнстом Хкелем из Йены.
Ранние антропологи придерживались точки зрения, что другие культуры были «примитивными» в эмбриологическом смысле, поскольку их развитие остановилось на уровне нашего собственного. Действительно, для определения такой культуры до сих пор используется слово «недоразвитая». Поскольку эволюция представляла собой последовательное добавление к вершине дерева, различные расы можно было расположить сверху вниз. (Действительно, их нужно было бы упорядочить линейно, поскольку это не модель с разветвленной цепью.) Ранее несколько историков науки упоминали, что Геккель был антисемитом и что его биология использовалась Третьим рейхом.Эти претензии были повторены на этом веб-сайте. Хотя его биология, безусловно, пыталась классифицировать человеческие группы по расовым характеристикам и использовалась для оправдания притязаний одной этнической группы на превосходство над другими, она не была явно антисемитской. Не был Геккель и антисемитом. Боб Ричардс (2007) обнаружил, что произошла путаница в идентичностях (среди прочего) с другим (и более молодым) Эрнстом Хкелем из Йены. В самом деле, в его рейтинге человечества семиты были достаточно хороши, выходя обычно чуть ниже арийцев.(Зато у чернокожих, финнов и ирландцев есть основания для претензий.)
В самом деле, в его рейтинге человечества семиты были достаточно хороши, выходя обычно чуть ниже арийцев.(Зато у чернокожих, финнов и ирландцев есть основания для претензий.)
Геккель привнес западное представление о Великой Цепи Бытия в эволюционную мысль. Подобно версиям Великой Цепи Средневековья, Ренессанса и Просвещения, она прославляла восхождение человека. А мужчина — это тот пол, который был важен. И расы, и полы ранжировались выше и ниже, и белые женщины, по сути, помещались на ту же ступеньку эволюционной лестницы, что и чернокожие мужчины или европейские младенцы. (Обратите внимание на последовательность: младшие стадии европейских самцов представлены взрослыми ниже по цепочке).Так, Карл Фогт, профессор естественной истории Женевского университета и современник Геккеля, утверждал (1864 г.): «Своей округлой вершиной и менее развитой задней долей негритянский мозг напоминает мозг наших детей, а выпуклостью теменной доли, у наших женщин». В заключение он заявил, что характеристики мозга вместе «отводят мозгу негра место рядом с мозгом белого ребенка». Считалось, что женщины тоже принадлежат к ним, поскольку Фогт также пришел к выводу, что «женский европейский череп гораздо больше похож на череп негра, чем на череп европейского мужчины.Фогт был не одинок. Он цитировал многочисленные исследования, в том числе исследование антрополога Хушке, который пришел к выводу, что «в негритянском мозгу как мозжечок, так и головной мозг, а также спинной мозг представлены женским и инфантильным европейским, а также Таким образом, чернокожие, женщины и дети связывают обезьян со взрослыми белыми самцами.
Считалось, что женщины тоже принадлежат к ним, поскольку Фогт также пришел к выводу, что «женский европейский череп гораздо больше похож на череп негра, чем на череп европейского мужчины.Фогт был не одинок. Он цитировал многочисленные исследования, в том числе исследование антрополога Хушке, который пришел к выводу, что «в негритянском мозгу как мозжечок, так и головной мозг, а также спинной мозг представлены женским и инфантильным европейским, а также Таким образом, чернокожие, женщины и дети связывают обезьян со взрослыми белыми самцами.
Можно увидеть это понятие линейной эволюции во многих социальных мысли. В религии это стало доминирующим взглядом на историю западной религиозной мысли: иудаизм воспроизвел язычество, а затем превзошел его.Затем христианство повторило язычество, иудаизм, а затем превзошло его. (Этот тезис стал известен как тезис Велльхаузена по имени человека, который наиболее четко его сформулировал). Таким образом, иудаизм рассматривался как зачаток христианства, более примитивная форма мысли, роль которой заключалась в том, чтобы подготовить мир к зрелой форме. В более истинно эволюционном смысле можно увидеть разветвленную модель религиозной мысли, в которой христианство и современный иудаизм возникли из иудаизма 2000 лет назад.
В более истинно эволюционном смысле можно увидеть разветвленную модель религиозной мысли, в которой христианство и современный иудаизм возникли из иудаизма 2000 лет назад.
В конце концов, Биогенетический Закон стал научно несостоятельным.(Восстание против этого «закона» было начато в середине 1890-х годов британским эмбриологом Адамом Седжвиком, который заметил накопление исключений из этого «правила» и смог переинтерпретировать более ранние результаты, не прибегая к нему. Закон стал союзником понятия все более подозрительного понятия наследования приобретенных признаков.К 1922 году Уолтер Гарстанг смог предоставить более сложный анализ взаимосвязи между эволюцией и развитием, показав, что изменения в развитии могут вызывать эволюционные изменения.) Однако и по сей день он остается популярным в обществе способом думать об эволюции. Доктор Спок (один из самых популярных либеральных мыслителей в Соединенных Штатах) использовал его в 1968 году для обсуждения развития человеческого плода, а реклама Continental Bank в Newsweek в 1980-х показывает линейный путь от протиста до банкира.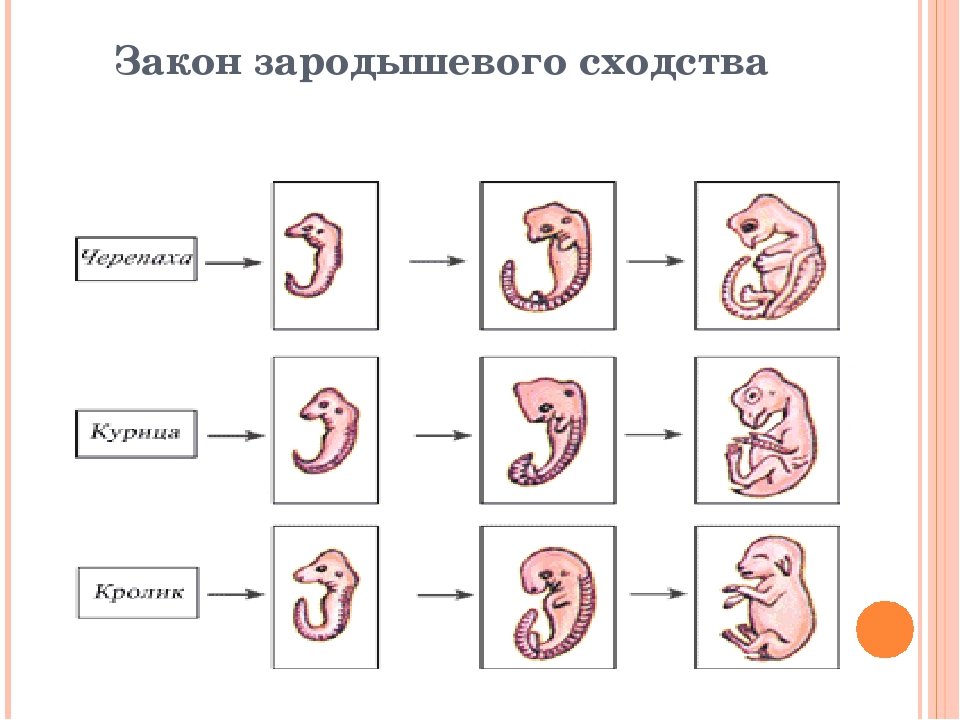 (белый, мужчина, с портфелем), когда банк утверждает, что превратился в «бескомпромиссный, полнокровный, не отвлекающийся, целеустремленный банк для бизнеса.«Мы должны понимать, что такие изображения эволюции все еще широко распространены в массовой культуре и что они способны причинить огромный вред».
(белый, мужчина, с портфелем), когда банк утверждает, что превратился в «бескомпромиссный, полнокровный, не отвлекающийся, целеустремленный банк для бизнеса.«Мы должны понимать, что такие изображения эволюции все еще широко распространены в массовой культуре и что они способны причинить огромный вред».
Цитированная литература
Garstang, W. 1922. Теория перепросмотра: критическое переформулирование биогенетического закона. Дж. Линн. соц. Зоол. 35: 81-101.
Гассман, Д. 1971. Научные истоки национал-социализма: социальный дарвинизм у Эрнста Геккеля и Немецкая монистическая лига. Макдональд, Лондон.
Gould, 1977. Онтогенез и филогения. Издательство Гарвардского университета, Кембридж.
Гулд, С. Дж. 1977. С тех пор, как Дарвин. Нортон, Нью-Йорк.
Haeckel, E. 1867. Generelle Morphologie der Organismen. Георг Раймер, Берлин.
Геккель, Э. 1879. Антропогения. Третье издание.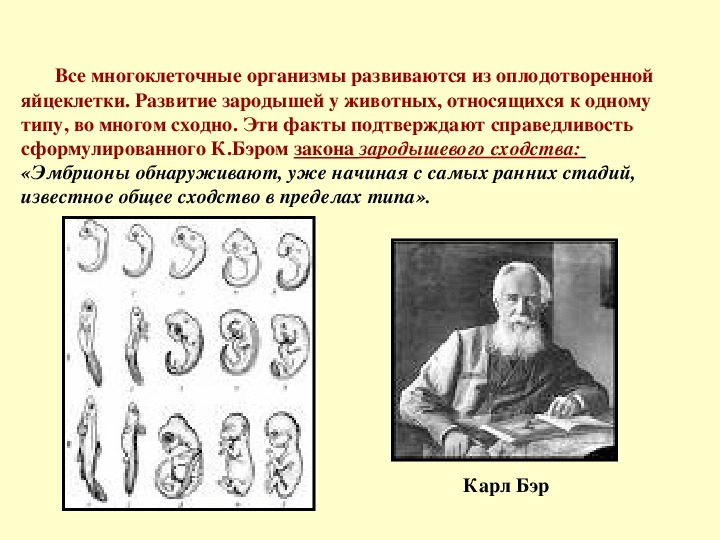 В. Энгельманн, Лейпциг.
В. Энгельманн, Лейпциг.
Геккель, Э. 1902. Загадка Вселенной. Harper and Brothers, Нью-Йорк.
Ричардс, Р.J. 2007. Предполагаемый антисемитизм Эрнста Геккеля и его вклад в нацистскую биологию. Биологическая теория 2: 97–103.
Спок, Б. 1968. Ребенок и уход за детьми. Исправленное издание. Карманные книги, Нью-Йорк.
Vogt, C. 1864. Лекции о человеке. Лонгман, Грин, Лонгман и Робертс, Лондон. п. 172-183.
von Baer, KE 1828. Entwickelungsgeschichte der Thiere: Beobachtung und Reflexion. Борнтргер, Конингсберг.
Преформизм — обзор | ScienceDirect Topics
Смена парадигм наследственности и эволюции
В конце 18 века врач и философ Эразм Дарвин (дедушка Чарльза Дарвина) дал импульс, на котором сформировалась основа современных теорий эволюции и наследственности.В своей основополагающей работе Э. Дарвин яростно отверг преформизм (теорию о том, что все организмы произошли от миниатюрных форм самих себя) и взамен изложил аристотелевскую теорию эпигенеза (идею о том, что организмы достигли окончательной взрослой формы, развиваясь из более простая, преждевременная форма) для синтеза последовательной теории эволюции. Основные элементы теории Э. Дарвина можно резюмировать в следующем отрывке из Zoonomia [1]:
Основные элементы теории Э. Дарвина можно резюмировать в следующем отрывке из Zoonomia [1]:
«Было бы слишком смело вообразить, что за большой промежуток времени, с тех пор как Земля начала существовать, возможно, за миллионы веков до начала истории человечества, было бы слишком смело воображать, что все теплокровные животные произошли от одной живой нити, которую ВЕЛИКОЕ ПРИЧИНО наделило животностью, способностью приобретать новые части, сопровождаться с новыми наклонностями, направляемыми раздражениями, ощущениями, волями и ассоциациями; и, таким образом, обладая способностью продолжать совершенствоваться благодаря своей собственной присущей ему деятельности и передавать эти улучшения через поколение своему потомству, миру без конца!»
Тринадцать лет спустя французский естествоиспытатель Жан-Батист Ламарк (1744–1829) близко поддержал мысли Э.Дарвин в своей обширной теории эволюции и наследственности опубликовал в виде трех основных работ: Recherches sur l’organisation des corps vivants (1802 г. ), Philosophie zoologique (1809 г.) и Histoire naturelle des animaux sans vertèbres (в семи томах). , 1815–1822). Ламарк постулировал, что взаимодействие человека с окружающей средой служит решающей движущей силой эволюции видов в направлении большей сложности и совершенства. Он считал, что эволюция происходила в виде постепенного, органичного процесса, в ходе которого взаимодействие индивида с окружающей средой вызывает отек органа тела за счет движения телесных жидкостей, и этот отек передается следующему поколению, где затем он еще больше улучшается за счет движения телесных жидкостей. тот же механизм.
), Philosophie zoologique (1809 г.) и Histoire naturelle des animaux sans vertèbres (в семи томах). , 1815–1822). Ламарк постулировал, что взаимодействие человека с окружающей средой служит решающей движущей силой эволюции видов в направлении большей сложности и совершенства. Он считал, что эволюция происходила в виде постепенного, органичного процесса, в ходе которого взаимодействие индивида с окружающей средой вызывает отек органа тела за счет движения телесных жидкостей, и этот отек передается следующему поколению, где затем он еще больше улучшается за счет движения телесных жидкостей. тот же механизм.
Согласно Ламарку, двумя главными движущими силами эволюции являются le pouvoir de vie (сила возрастающей сложности) и l’influence des circonstances (сила, заставляющая организмы приспосабливаться к окружающей среде). Что касается l’influence des circonstances , Ламарк выделил два естественных закона, которые действуют во всех организмах: (1) в результате использования и неиспользования индивидуумы приобретают или теряют характеристики в течение своей жизни и (2) характеристики, приобретенные в течение жизни индивидуума, наследуются. от одного поколения к другому.Эти два принципа, теперь вместе известные как теория наследования приобретенных признаков, не были уникальными для Ламарка, и эта идея не была центральной в ламарковской теории эволюции; однако Ламарк был неразрывно связан с этой концепцией [2]. По мере углубления понимания экологически индуцированных негенетических путей наследования признаков ламаркизм в этом отношении, по-видимому, возродился, но не без противоречий [3–10]. Таким образом, здесь мы сосредоточимся на этом конкретном аспекте теории Ламарка и на том, как она впала в немилость, а затем вновь обрела легитимность в качестве краеугольного камня трансгенерационной эпигенетики в 21 веке.
от одного поколения к другому.Эти два принципа, теперь вместе известные как теория наследования приобретенных признаков, не были уникальными для Ламарка, и эта идея не была центральной в ламарковской теории эволюции; однако Ламарк был неразрывно связан с этой концепцией [2]. По мере углубления понимания экологически индуцированных негенетических путей наследования признаков ламаркизм в этом отношении, по-видимому, возродился, но не без противоречий [3–10]. Таким образом, здесь мы сосредоточимся на этом конкретном аспекте теории Ламарка и на том, как она впала в немилость, а затем вновь обрела легитимность в качестве краеугольного камня трансгенерационной эпигенетики в 21 веке.
Теория эволюции Ламарка прочно утвердилась, когда два английских естествоиспытателя, Чарльз Роберт Дарвин (1809–1882) и Альфред Рассел Уоллес (1823–1913), независимо друг от друга пришли к одному и тому же выводу, что процесс естественного отбора является основным механизмом эволюции. эволюция. Ч. Дарвин, в частности, был сторонником общей теории эволюции Ламарка и по большей части принимал ламарковский принцип использования/неиспользования как законный механизм эволюции. В поддержку теории Ламарка Дарвин предложил дополнительную гипотезу, которую он назвал «Пангенезис» в своей книге « Изменчивость растений и животных при доместикации » [11].Дарвин предположил, что сигналы окружающей среды могут побуждать соматические клетки выделять микроскопические «геммулы» или «пангены», которые циркулируют в крови и в конечном итоге достигают половых желез, накапливаясь в зародышевых клетках. Геммулы, по сути, представляют собой способ передачи информации от соматических клеток к зародышевым, влияя таким образом на фенотип следующего поколения. В то время пангенезис был в значительной степени отвергнут научным сообществом, поскольку не было эмпирических доказательств существования пангенов, а сэр Фрэнсис Гальтон (1822–1911) продемонстрировал, что переливание крови между кроликами разного цвета меха не приводит к изменению цвет меха в потомстве.Несмотря на необоснованную гипотезу в то время, дарвиновские пангены очень напоминают то, что мы знаем в настоящее время о способах наследования, не основанных на ДНК (т.
В поддержку теории Ламарка Дарвин предложил дополнительную гипотезу, которую он назвал «Пангенезис» в своей книге « Изменчивость растений и животных при доместикации » [11].Дарвин предположил, что сигналы окружающей среды могут побуждать соматические клетки выделять микроскопические «геммулы» или «пангены», которые циркулируют в крови и в конечном итоге достигают половых желез, накапливаясь в зародышевых клетках. Геммулы, по сути, представляют собой способ передачи информации от соматических клеток к зародышевым, влияя таким образом на фенотип следующего поколения. В то время пангенезис был в значительной степени отвергнут научным сообществом, поскольку не было эмпирических доказательств существования пангенов, а сэр Фрэнсис Гальтон (1822–1911) продемонстрировал, что переливание крови между кроликами разного цвета меха не приводит к изменению цвет меха в потомстве.Несмотря на необоснованную гипотезу в то время, дарвиновские пангены очень напоминают то, что мы знаем в настоящее время о способах наследования, не основанных на ДНК (т.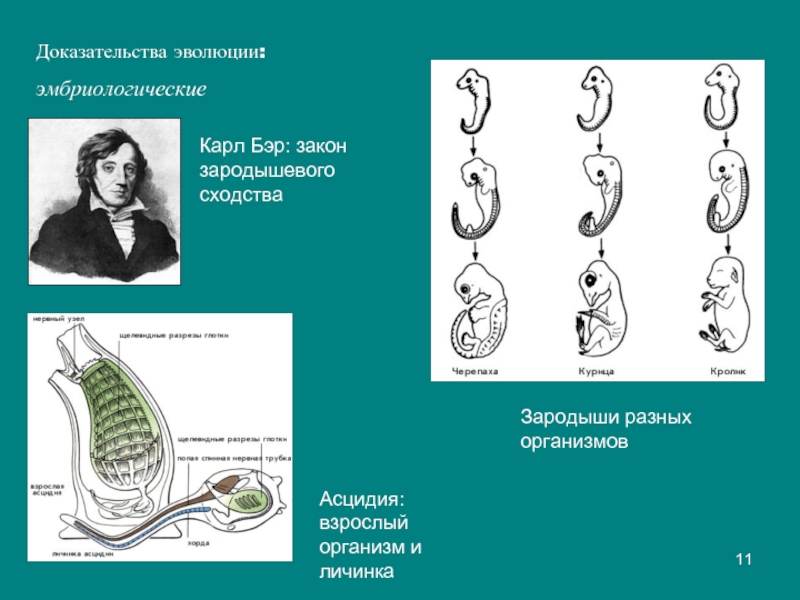 е. прионы, некодирующие РНК и экзосомы) [12–17]. На самом деле, появляется все больше доказательств того, что переливание крови может вызывать наследственные изменения у животных, специфичные для заболевания, вероятно, за счет индукции модификаций хроматина путем доставки циркулирующих РНК в сперму [18,19].
е. прионы, некодирующие РНК и экзосомы) [12–17]. На самом деле, появляется все больше доказательств того, что переливание крови может вызывать наследственные изменения у животных, специфичные для заболевания, вероятно, за счет индукции модификаций хроматина путем доставки циркулирующих РНК в сперму [18,19].
На рубеже веков мощные реакционные силы привели к отказу от ламаркистской эволюции.Немецкий биолог Август Вейсманн (1834–1914), вероятно, был самым влиятельным критиком ламарковской наследственности. В своей теории зародышевой плазмы Вейсманн установил резкое различие между функциями соматических клеток, заявив, что зародышевые клетки являются единственными проводниками наследственности, тогда как соматические клетки отвечают за функции организма [20,21]. Он решительно отверг ламарковскую теорию эволюции использования/неиспользования в том, что должно было быть известно как барьер Вейсмана. Согласно Вейсману, зародышевые клетки невосприимчивы к воздействиям окружающей среды и соматическим воздействиям, и, таким образом, приобретенные признаки не могут быть унаследованы будущими поколениями. В поддержку своей теории однонаправленного потока информации от зародышевых клеток к соматическим Вейсманн продемонстрировал, что приобретенный при жизни признак — искусственное укорочение мышиного хвоста — не может быть унаследован последующими поколениями [20]. Хотя эксперимент имел недостаток в том, что он не проверял наследование биологически значимого признака, который Ламарк счел бы важным в эволюции вида, он бросил огромную тень сомнения на ламарковскую эволюцию.Сегодня мы знаем, что изменения окружающей среды действительно могут влиять на гаметы посредством регуляции соматического фенотипа и наоборот [8,16,22–25].
В поддержку своей теории однонаправленного потока информации от зародышевых клеток к соматическим Вейсманн продемонстрировал, что приобретенный при жизни признак — искусственное укорочение мышиного хвоста — не может быть унаследован последующими поколениями [20]. Хотя эксперимент имел недостаток в том, что он не проверял наследование биологически значимого признака, который Ламарк счел бы важным в эволюции вида, он бросил огромную тень сомнения на ламарковскую эволюцию.Сегодня мы знаем, что изменения окружающей среды действительно могут влиять на гаметы посредством регуляции соматического фенотипа и наоборот [8,16,22–25].
В ответ на немедленную и продолжающуюся оппозицию сторонники ламаркистской эволюции поспешили продемонстрировать наследование приобретенных признаков. Многие исследования были ошибочными, не вызывали доверия, были омрачены политическими программами того времени или вообще не соответствовали ламаркистскому духу. Среди первых сторонников ламаркистского наследования были Шарль-Эдуар Браун-Секар, Гастон Боннье и Фредерик Меррифилд. В их попытках доказать обоснованность теорий Ламарка отсутствовали эмпирические доказательства реальных механизмов наследственности, и поэтому они подверглись критике со стороны ведущих генетиков. Заметным недостатком теории Ламарка была фабрикация Трофимом Денисовичем Лысенко (1898–1976) экспериментов для поддержки негенетического наследования признаков в 1930-х и 1940-х годах [26]. Лысенко, русский агроном, отстаивал идеи Ламарка и пытался применить ламарковские теории для продвижения и улучшения существующих методов ведения сельского хозяйства.Отрицание Лысенко существования генов и концепции эволюции путем естественного отбора во многом проистекало из его презрения к тому, что он считал западной наукой. Его повторное открытие «яровизации», позже переведенной как «яровизация», показало, что предварительное воздействие низких температур на семена увеличивает цветение растений и впоследствии повышает урожайность. Чтобы поддержать свои идеи, которые стали известны как лысенковщина, Лысенко представил ложные доказательства ламарковской наследственности яровизации.
В их попытках доказать обоснованность теорий Ламарка отсутствовали эмпирические доказательства реальных механизмов наследственности, и поэтому они подверглись критике со стороны ведущих генетиков. Заметным недостатком теории Ламарка была фабрикация Трофимом Денисовичем Лысенко (1898–1976) экспериментов для поддержки негенетического наследования признаков в 1930-х и 1940-х годах [26]. Лысенко, русский агроном, отстаивал идеи Ламарка и пытался применить ламарковские теории для продвижения и улучшения существующих методов ведения сельского хозяйства.Отрицание Лысенко существования генов и концепции эволюции путем естественного отбора во многом проистекало из его презрения к тому, что он считал западной наукой. Его повторное открытие «яровизации», позже переведенной как «яровизация», показало, что предварительное воздействие низких температур на семена увеличивает цветение растений и впоследствии повышает урожайность. Чтобы поддержать свои идеи, которые стали известны как лысенковщина, Лысенко представил ложные доказательства ламарковской наследственности яровизации. Точно так же его сторонники утверждали, что открыли негенетические способы заставить рожь стать пшеницей. К сожалению, Иосиф Сталин, а также многие другие лидеры бывшего Советского Союза и Китая в то время решительно поддержали необоснованные утверждения Лысенко и обещание сельскохозяйственной революции и приказали заключить в тюрьму и казнить тысячи генетиков, выступавших против лысенковщины.
Точно так же его сторонники утверждали, что открыли негенетические способы заставить рожь стать пшеницей. К сожалению, Иосиф Сталин, а также многие другие лидеры бывшего Советского Союза и Китая в то время решительно поддержали необоснованные утверждения Лысенко и обещание сельскохозяйственной революции и приказали заключить в тюрьму и казнить тысячи генетиков, выступавших против лысенковщины.
В то время как лысенковщина и ламаркизм утвердились в Восточном блоке, в остальном мире началась эра геноцентрической науки, что еще больше подорвало доверие к ламаркизму и теории пангенезиса Дарвина.Это движение, начавшееся с середины 1930-х до конца 1940-х годов и продолжавшееся вплоть до 1970-х годов, известно как «Современный синтез» (или неодарвинизм) [27]. К Современному синтезу привело открытие хромосом Вальтером Флеммингом (1843–1905) в 1879 году и растущее количество свидетельств того, что гены являются единицами наследственности, благодаря работам Эдмунда Уилсона (1856–1939), Теодора Бовери (1862–1915). , Уолтер Саттон (1877–1916) и Томас Х. Морган (1866–1944). Именно повторное открытие работы Грегора Менделя о прерывистом наследовании признаков у растений гороха стало ключом к открытию этой новой эры.В 1866 году Мендель опубликовал свои основные выводы о механизмах наследования признаков, теперь известные как закон наследования Менделя (т. Е. Закон сегрегации и закон независимого ассортимента). В то время работа Менделя в значительной степени игнорировалась из-за популярности теории смешанного наследования, согласно которой черты, наблюдаемые у потомства, являются результатом усреднения или смешения родительских черт. На рубеже 20-го века Хьюго де Врис, Карл Корренс, Эрих фон Чермак и Уильям Джаспер Спиллман, все из которых не знали о результатах работы Менделя, независимо друг от друга провели эксперименты, которые подтвердили и расширили менделевские законы сегрегации и независимых ассортимент.В это время Уильям Бейтсон, активный сторонник менделевского наследования, ввел термин «генетика» для описания изучения того, как наследование признаков от родителей к потомству может привести к множеству вариаций признаков.
, Уолтер Саттон (1877–1916) и Томас Х. Морган (1866–1944). Именно повторное открытие работы Грегора Менделя о прерывистом наследовании признаков у растений гороха стало ключом к открытию этой новой эры.В 1866 году Мендель опубликовал свои основные выводы о механизмах наследования признаков, теперь известные как закон наследования Менделя (т. Е. Закон сегрегации и закон независимого ассортимента). В то время работа Менделя в значительной степени игнорировалась из-за популярности теории смешанного наследования, согласно которой черты, наблюдаемые у потомства, являются результатом усреднения или смешения родительских черт. На рубеже 20-го века Хьюго де Врис, Карл Корренс, Эрих фон Чермак и Уильям Джаспер Спиллман, все из которых не знали о результатах работы Менделя, независимо друг от друга провели эксперименты, которые подтвердили и расширили менделевские законы сегрегации и независимых ассортимент.В это время Уильям Бейтсон, активный сторонник менделевского наследования, ввел термин «генетика» для описания изучения того, как наследование признаков от родителей к потомству может привести к множеству вариаций признаков. Другими участниками этого проявления силы были Рональд Фишер, Сьюэлл Райт и Дж. Б. Холдейн, а также их систематическое изучение популяционной генетики и количественной генетики. Это побудило других, таких как Феодосий Добржанский и Томас Х. Морган, продолжить исследования, которые провозгласили идею о том, что генетика является единственной основой дарвиновской теории естественного отбора.Важно отметить, что во времена Современного синтеза большинство ученых отвергли идею эволюции через приобретенные признаки и сосредоточили изучение наследственности исключительно на генах. Это чувство было незыблемым более 40 лет после его зарождения. Только в последние несколько десятилетий 20-го века, с открытием наследуемых эпигенетических состояний, ламарковское наследование начало сбрасывать покров лженауки и стало рассматриваться как законная парадигма эволюции, объединяющая генетические и негенетические компоненты признака. наследственность.
Другими участниками этого проявления силы были Рональд Фишер, Сьюэлл Райт и Дж. Б. Холдейн, а также их систематическое изучение популяционной генетики и количественной генетики. Это побудило других, таких как Феодосий Добржанский и Томас Х. Морган, продолжить исследования, которые провозгласили идею о том, что генетика является единственной основой дарвиновской теории естественного отбора.Важно отметить, что во времена Современного синтеза большинство ученых отвергли идею эволюции через приобретенные признаки и сосредоточили изучение наследственности исключительно на генах. Это чувство было незыблемым более 40 лет после его зарождения. Только в последние несколько десятилетий 20-го века, с открытием наследуемых эпигенетических состояний, ламарковское наследование начало сбрасывать покров лженауки и стало рассматриваться как законная парадигма эволюции, объединяющая генетические и негенетические компоненты признака. наследственность.
От открытий Эрнста Геккеля до современных взглядов
Брюнет, Т. , Ларсон, Б.Т., Линден, Т.А., Вермейдж, М.Дж., Макдональд, К., и
, Ларсон, Б.Т., Линден, Т.А., Вермейдж, М.Дж., Макдональд, К., и
Кинг, Н. (2019). Коллективная сократимость, регулируемая светом, у многоклеточных хоанофлагеллят
. Наука, 366 (6463), 326–334.
Burkhardt, P., Stegmann, C.M., Cooper, B., Kloepper, TH, Imig, C.,
Varoqueaux, F., Wahl, M.C., & Fassauer, D. (2011). Первичный
нейросекреторный аппарат выявлен у хоанофлагеллят
Monosiga brevicollis.Труды Национальной академии наук
Соединенных Штатов Америки, 108 (37), 15264–15269.
Дарвин, К. (1859 г.). Происхождение видов путем естественного отбора или сохранение привилегированных рас в жизненной борьбе. Дж. Мюррей.
Дайел, М.Дж., Алегадо, Р.А., Фэйрклаф, С.Р., Левин, Т.С., Николс, С.А.,
Макдональд, К., и Кинг, Н. (2011). Дифференцировка клеток и
морфогенез у колониеобразующей хоанофлагелляты Salpingoeca
rosetta.Биология развития, 357 (1), 73–82.
Домазет-Лошо, Т., и Таутц, Д. (2010). Филогенетически основанный индекс возраста транскриптома
отражает образцы онтогенетической дивергенции.
Природа, 468, 815–819.
Дюбул, Д. (1994). Временная коллинеарность и филотипическая прогрессия:
Основа стабильности бауплана позвоночных и эволюция
морфологий посредством гетерохронии. Развитие, 135–142.
Fairclough, S.R., Chen, Z.,Kramer,E.,Zeng,Q.,Young,S.,
Robertson, HM, Begovic, E., Richter, DJ, Russ, C.,
Westbrook,MJ,Manning,G.,Lang,BF , Хаас Б., Нусбаум К.,
и Кинг Н. (2013). Эволюция генома предметазоя и
регуляция дифференцировки клеток у хоанофлагеллят
Salpingoeca rosetta. Genome Biology, 14(2), R15.
Гёде Р., Науманн Б., Лаундон Д., Имиг К., Макдональд К.Л.,
Купер Б.Х. и Буркхардт П. (2020).Хоанофлагелляты и
предки нейросекреторных пузырьков. bioRxiv.https://doi.org/10.
1101/2020.05.24.111997
Гулд, С. Дж. (1977). Онтогенез и филогенез. Belknap Press of Harvard
University Press.
Геккель, Э. (1866 г.). Общая морфология организмов (2 Bde). Г.
Г.
Реймер.
Геккель, Э. (1868 г.). Natürliche Schöpfungsgeschichte. Г. Реймер.
Геккель, Э. (1870a). Biologische Studien: Studien über Moneren und andere
Protisten: nebst einer Rede über Entwickelungsgang und Aufgabe der
Zoologie (Vol.1). Энгельманн.
Геккель, Э. (1870b). Die Catallacten, eine neue Protisten-Gruppe: (Abdruck
ad «Jenaischen Zeitschrift». Bd. VI, Hft. 1.). Густав Фишер.
Геккель, Э. (1872 г.). Monographie der Kalkschwämme (3 Bde). Г. Реймер.
Геккель, Э. (1874a). Anthropogenie oder Entwickelungsgeschichte des
Menschen (1-е изд.). Вильгельм Энгельманн.
Геккель, Э. (1874b). Die Gastrea-Theorie, die phylogenetische Klassifikation
des Tierreichs, und die Homologie der Keimblätter.Jenaische Zeitschrift
für Naturwissenschaften, 8,1–55.
Геккель, Э. (1875 г.). Die Gastrula und die Eifurchung der Thiere. Jenaische
Zeitschr. ф. Натургеш. IX. Бд.
Геккель, Э.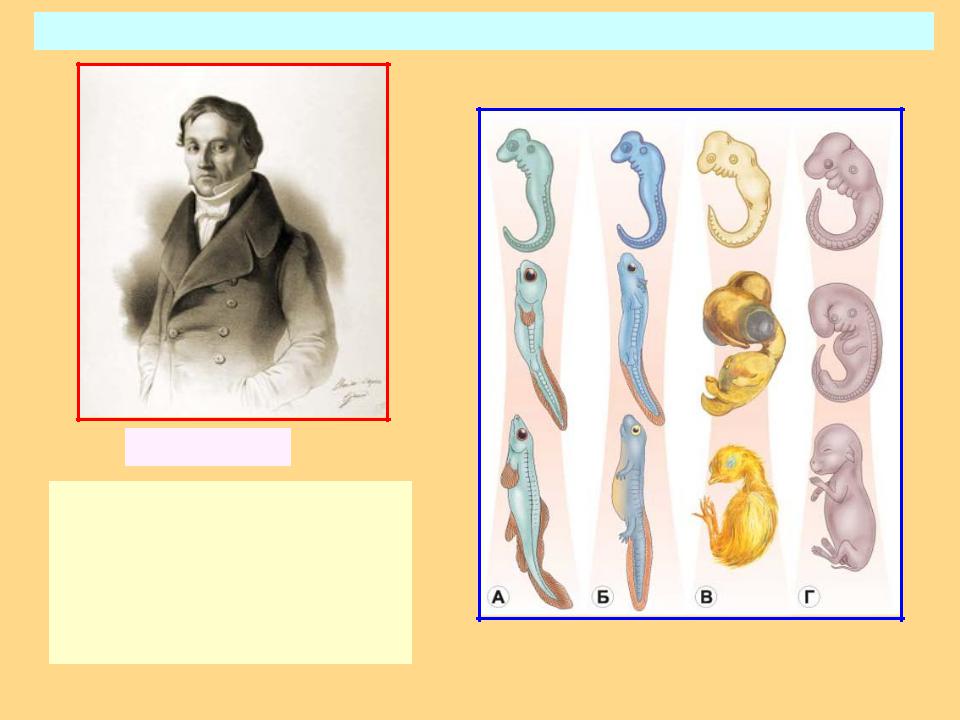 (1877a). Nachträge zur Gastrea-Theorie. Jenaische Zeitschrift
(1877a). Nachträge zur Gastrea-Theorie. Jenaische Zeitschrift
für Naturwissenschaft, herausgegeben von der Medicinisch-
Naturwissenschaftlichen Gesellschaft zu Jena, 11, 55–98.
Геккель, Э. (1877b). Studien zur Gastraea-theorie. Герман Даффт.
Геккель, Э. (1879 г.). Natürliche Schöpfungsgeschichte (7-е изд.). Г. Реймер.
Геккель, Э. (1909). Anthropogenie oder Entwickelungsgeschichte des Mensch
(6-е изд.). Вильгельм Энгельманн.
Холл, Б.К. (1997). Филотипическая стадия или фантом: Существует ли высококонсервативная эмбриональная стадия у позвоночных? Тенденции в экологии и
Эволюция, 12, 461–463.
Холл, Б.К. (2012). Эволюционная биология развития. Springer Science &
Business Media.
Хазкани-Ково, Э., Шерсть, Д., и Граур, Д. (2005). В поисках филотипической стадии
позвоночных: молекулярное исследование модели развития песочных часов
и третьего закона фон Бэра. Журнал
экспериментальной зоологии, часть B: молекулярная эволюция и развитие,
304B, 150–158.
Хоссфельд, У., Левит, Г.С., и Кучера, У. (2019). Эрнст Геккель
(1843–1919): немецкий Дарвин и его влияние на современную
биологию.Специальный выпуск. Теория биологических наук, 138 (1), 1–202.
Хосфельд, У., и Олссон, Л. (2003). Дорога из Геккеля. Йенская
традиция в эволюционной морфологии и происхождение «Эво-Дево».
Биология и философия, 18(2), 285–307.
Ири, Н., и Куратани, С. (2011). Сравнительный транскриптомный анализ
выявляет филотипический период позвоночных во время органогенеза.
Природные коммуникации, 2,1–8.
Ириэ, Н., и Сехара-Фудзисава, А.(2007). Филотипическая стадия позвоночных и
ранняя билатеральная стадия в эмбриогенезе мышей, определяемая
геномной информацией. BMC Biology, 5,1.https://doi.org/10.1186/
1741-7007-5-1
Junker, T., & Hoßfeld, U. (2009). Die Entdeckung der Evolution: Eine
Revolutionäre Theorie und ihre Geschichte. Wissenschaftliche
Buchgesellschaft.
Калинка А.Т., Варга К.М., Джеррард Д.Т., Прейбиш С., Коркоран Д.L.,
Джарреллс, Дж., Олер, У., Бергман, К.М., и Томанчак, П. (2010). Дивергенция экспрессии гена
резюмирует модель развития песочных часов
. Природа, 468, 811–816.
Кинг, Н., Уэстбрук, М.Дж., Янг, С.Л., Куо, А., Абедин, М., Чепмен, Дж.,
Фэйркло, С., Хеллстен, У., Исогай, Ю., Летуник, И. ., Марр М., Пинкус Д.,
Патнэм Н., Рокас А., Райт К.Дж., Зузов Р., Диркс В., Гуд М.,
Гудстейн Д., … Рохсар, Д. (2008). Геном
хоанофлагеллят Monosiga brevicollis и происхождение многоклеточных животных.
Природа, 451(7180), 783–788.
Колчинский Э. и Левит Г. С. (2019). Рецепция Геккеля в дореволюционной России. Теория биологических наук, 138(1), 73–88.
Ковалевский А. О. (1866). Entwicklungsgeschichte дер einfachen Ascidien.
Мем. де л’Акад. Имп. де наук де Санкт‐Петербург. сер. VII. TX, № 15.
Ларсон, Б. Т., Руис-Эрреро, Т., Ли, С. , Кумар, С., Махадеван, Л., и Кинг, Н.
, Кумар, С., Махадеван, Л., и Кинг, Н.
(2020). Биофизические принципы самоорганизации хоанофлагеллят.
Proceedings of the National Academy of Sciences of the United States of
America,117, 1303–1311.
Ледбитер, BSC (1983). История жизни и ультраструктура новых
морских видов Proterospongia (Choanoflagellida). Журнал Морской биологической ассоциации
Соединенного Королевства, 63, 135–160.
Ледбитер, BSC (2015). Хоанофлагелляты. Кембриджский университет
Press.
Левин М., Хашимшони Т., Вагнер, Ф., и Янаи, И. (2012). Вехи развития
акцентируют экспрессию генов в эмбрионе Caenorhabditis.
Ячейка развития, 22, 1101–1108.
Левит, Г. С. (2007). Корни Evo-Devo в России: существует ли
характеристика «русской традиции»? Theory in Biosciences, 4, 131–148.
Левит, Г.С., Хоссфельд, У., и Олссон, Л. (2015). Алексей Северцов и Адольф
Наеф: пересмотр биогенетического закона Геккеля. История и философия
История и философия
наук о жизни, 36 (3), 357–370.
Михайлов К.В., Константинова А.В., Никитин М.А., Трошин П.В.,
Русин Л.Ю., Любецкий В.А., Панчин Ю.В., Мыльников А.П.,
Мороз Л.Л., Кумар С., Алеошин, ВВ (2009). Происхождение
Metazoa: переход от временной к пространственной дифференцировке клеток.
BioEssays, 31, 758–768.
Мюллер, Ф. (1864 г.). Для Дарвина. Вильгельм Энгельманн.
Олссон, Л., и Хосфельд, У. (2007). Die Entwicklung: Die Zeit des Lebens.
Ausgewählte Themen zur Geschichte der Entwicklungsbiologie. В E.
Höxtermann, & H. Hilger (Eds.), Lebenswissen: Eine Einführung in die
Geschichte der Biologie (стр. 218–243). Природа и текст.
Олссон, Л., Левит, Г.С., и Хоссфельд, У. (2010). Эволюционная биология развития
: ее концепции и история с акцентом на русский и
немецкий вклад. Naturwissenschaften, 97 (11), 951–969.
Олссон Л., Левит Г.С. и Хоссфельд, У. (2017). «Биогенетический закон» в
«Биогенетический закон» в
зоологии: от формулировки Эрнста Геккеля до современных подходов.
Теория биологических наук, 136 (1–2), 19–29.
Piesecka, B., Lichocki, P., Moretti, S., Bergmann, S., & Robinson-Rechavi,
M. (2013). Песочные часы и модели ранней консервации:
26
|
ЛЕВИТ И ДР.
Филотипическая стадия позвоночных и ранняя стадия, связанная с билатериями, в эмбриогенезе мышей, определяемая геномной информацией | BMC Biology
Оценка наследственной природы эмбриогенеза мыши
На основании того, что филотипическая стадия позвоночных должна выражать наибольшее соотношение генов, которые предположительно существовали у предковых позвоночных, или генов позвоночных (см. Методы и Рисунок 1 для определения генов позвоночных , также см. Дополнительный файл 1: Таксономическая классификация гомологов мышиных белок-кодирующих генов по таксономическому диапазону.) мы установили следующий индекс:
Рисунок 1
Наборы данных генома, используемые для определения эволюционных классификаций . Эволюционные классификации, определенные в нашем анализе, указаны вместе со стрелками, покрывающими диапазон их предполагаемого эволюционного происхождения (например, позвоночных гена представляют гены, которые, как считается, уже существовали до эволюции позвоночных). Цифры в скобках указывают на гены развития и общее количество генов, классифицированных в каждой эволюционной классификации.Эти классификации согласуются с недавним сообщением [25, 26] о том, что большинство дупликаций генов развития произошло до эволюции билатеральных и позвоночных, что подтверждает правомерность нашего определения гомологов (см. Дополнительный файл 1: Таксономическая классификация гомологов мышиных белков, кодирующих гены в соответствии с таксономическим диапазоном.). См. также Методы определения эволюционных классификаций и «генов развития».
Эволюционные классификации, определенные в нашем анализе, указаны вместе со стрелками, покрывающими диапазон их предполагаемого эволюционного происхождения (например, позвоночных гена представляют гены, которые, как считается, уже существовали до эволюции позвоночных). Цифры в скобках указывают на гены развития и общее количество генов, классифицированных в каждой эволюционной классификации.Эти классификации согласуются с недавним сообщением [25, 26] о том, что большинство дупликаций генов развития произошло до эволюции билатеральных и позвоночных, что подтверждает правомерность нашего определения гомологов (см. Дополнительный файл 1: Таксономическая классификация гомологов мышиных белков, кодирующих гены в соответствии с таксономическим диапазоном.). См. также Методы определения эволюционных классификаций и «генов развития».
Индекс предков позвоночных на стадии развития k = V k /N k
Где V k : количество неизбыточных генов позвоночных , экспрессированных на стадии k; N k : общее количество неизбыточных генов, экспрессированных на стадии k.
Аналогично, анализы с генами Bilaterian , Chordate , Tetrapod и Amniote также выявили бы характер каждой эмбриональной стадии в таксономическом контексте (см. Методы определения консервативных генов).
Для анализа индексов предков эмбриональных стадий мышей мы собрали наборы генов мыши из базы данных ENSEMBL [17] и профили экспрессии генов из базы данных Expressed Sequence Tags Database (dbEST) [18] и считали ген «экспрессированным». если соответствующий идентификатор UniGene [19] присутствовал в любой из классифицированных библиотек EST.Библиотеки EST, классифицированные по каждой стадии, перечислены в таблице 1. Кроме того, чтобы сделать этот анализ устойчивым к колебаниям во времени эмбриональных стадий, мы воспользовались преимуществами анализа подвижной группы (аналогичного хорошо известному методу «анализа скользящего среднего») для расчета индекс предка двух последовательных этапов:
Таблица 1 Идентификаторы библиотек EST, классифицированные по описанию этапов разработки. Каждая классифицированная стадия развития мыши содержала в среднем около 22 300 (409–71 504) отсчетов EST, что в среднем соответствовало 2900 (194–5 923) неповторяющихся записей гена ENSEMBL.Статус развития был основан на Манипулирование эмбрионом мыши ) [27] и Мышь [28].
Каждая классифицированная стадия развития мыши содержала в среднем около 22 300 (409–71 504) отсчетов EST, что в среднем соответствовало 2900 (194–5 923) неповторяющихся записей гена ENSEMBL.Статус развития был основан на Манипулирование эмбрионом мыши ) [27] и Мышь [28].
Индекс предков позвоночных на сгруппированной стадии k = V k , k+1 /N k , k+1
экспрессируется на стадии k или k+1; N k , k+1 : общее количество неизбыточных генов, экспрессированных на стадии k или k+1.
С помощью этих методов мы рассчитали индексы предков билатеральных, хордовых, позвоночных, четвероногих и амниот (рис. 2А–Е).
Рисунок 2
Анализ движущихся групп индивидуальных индексов предков, полученных во время эмбриогенеза . A–E: анализ движущихся групп индексов предков во всех генах (25 613 генов). Номер сгруппированной стадии представляет более раннюю стадию пары стадий развития (например, экспрессия генов сгруппированной стадии 14 представляет неперекрывающиеся гены, экспрессированные на стадии 14 или 15; см. Таблицу 1 для определения стадии). Каждый этап содержал в среднем 22 744 отсчета EST (4 418–44 210), что соответствовало в среднем 4 396 (1 493–6 507) неизбыточных ENSEMBL M.musculus ген. F–G: анализ движущихся групп индексов предков в генах развития. Каждая стадия содержала в среднем 1479 отсчетов EST (282–3406), что соответствовало 303 (94–491) неизбыточным генам ENSEMBL M. musculus . E, M и L обозначают ранний, средний и поздний эмбриональные периоды, охватывая неизбыточные гены, экспрессируемые на стадиях 2–7, 11–18 и 25–31 соответственно. Статистический анализ проводили с помощью точного критерия Фишера. Значимые различия отмечены звездочками.нс: P > 0,05. На каждом графике относительные площади кружков отражают количество экспрессированных (всего или связанных с развитием) генов на каждой стадии.
Таблицу 1 для определения стадии). Каждый этап содержал в среднем 22 744 отсчета EST (4 418–44 210), что соответствовало в среднем 4 396 (1 493–6 507) неизбыточных ENSEMBL M.musculus ген. F–G: анализ движущихся групп индексов предков в генах развития. Каждая стадия содержала в среднем 1479 отсчетов EST (282–3406), что соответствовало 303 (94–491) неизбыточным генам ENSEMBL M. musculus . E, M и L обозначают ранний, средний и поздний эмбриональные периоды, охватывая неизбыточные гены, экспрессируемые на стадиях 2–7, 11–18 и 25–31 соответственно. Статистический анализ проводили с помощью точного критерия Фишера. Значимые различия отмечены звездочками.нс: P > 0,05. На каждом графике относительные площади кружков отражают количество экспрессированных (всего или связанных с развитием) генов на каждой стадии.
Первоначально нас удивило появление раннего пика вокруг сгруппированных стадий с 4 по 10 при анализе генов Bilaterian и Chordate (рис. 2A, B). Широко оцененный анализ также подтвердил эту тенденцию; как ранний (стадии 2–7), так и средний эмбриональные периоды (стадии 11–18) имеют значительно более высокие индексы предков, чем поздний период (стадии 25–31).Заметно более высокие индексы предков, показанные на рис. 2A, B, указывают на существование новых, хорошо сохранившихся стадий, предположительно несущих характеристики билатерий на стадиях от расщепления до бластулы в эмбриогенезе мышей.
2A, B). Широко оцененный анализ также подтвердил эту тенденцию; как ранний (стадии 2–7), так и средний эмбриональные периоды (стадии 11–18) имеют значительно более высокие индексы предков, чем поздний период (стадии 25–31).Заметно более высокие индексы предков, показанные на рис. 2A, B, указывают на существование новых, хорошо сохранившихся стадий, предположительно несущих характеристики билатерий на стадиях от расщепления до бластулы в эмбриогенезе мышей.
Филотипическая стадия позвоночных в эмбриогенезе мыши
Затем мы рассчитали индексы предков для поиска предполагаемой филотипической стадии эмбриогенеза мыши. В отличие от индексов предков билатеральных и хордовых, индексы предков позвоночных, четвероногих и амниот не показали заметных пиков (рис. 2C-E.Вместо этого мы получили слабо изогнутые графики с пиками вокруг сгруппированных стадий с 8 по 18 (рис. 2C–E), охватывающие предполагаемые периоды филотипической стадии позвоночных, предложенные сравнительными морфологическими исследованиями (например, стадия фарингулы [10], стадия ранней сегментации сомитов). [11] и стадию хвостовой почки [12]). Важно отметить, что индекс предков позвоночных в среднем эмбриональном периоде был значительно выше, чем в ранний и поздний эмбриональные периоды (рис. 2С), что также видно по индексам предков четвероногих и амниот (рис. 2D, E).Все эти кривые (рис. 2C-E) значительно отличались от полученных и показанных на рис. 2A, B (P <0,001; см. Методы оценки распределения индекса предков).
[11] и стадию хвостовой почки [12]). Важно отметить, что индекс предков позвоночных в среднем эмбриональном периоде был значительно выше, чем в ранний и поздний эмбриональные периоды (рис. 2С), что также видно по индексам предков четвероногих и амниот (рис. 2D, E).Все эти кривые (рис. 2C-E) значительно отличались от полученных и показанных на рис. 2A, B (P <0,001; см. Методы оценки распределения индекса предков).
Если бы мы рассматривали план тела и морфологический паттерн как показания генетических программ развития, то индексы предков, рассчитанные для генов развития, улучшили бы наше понимание сохранения стадий между 8 и 18. На этом предположении мы сосредоточились на генах развития мыши, определенных в базе данных Gene Ontology (см. Методы определения генов развития), чтобы получить индексы предков отдельных стадий.Самый высокий индекс предков наблюдался на сгруппированной стадии 14 (стадии 14–15) и был значительно выше, чем значения как раннего, так и позднего эмбрионального периода (рис.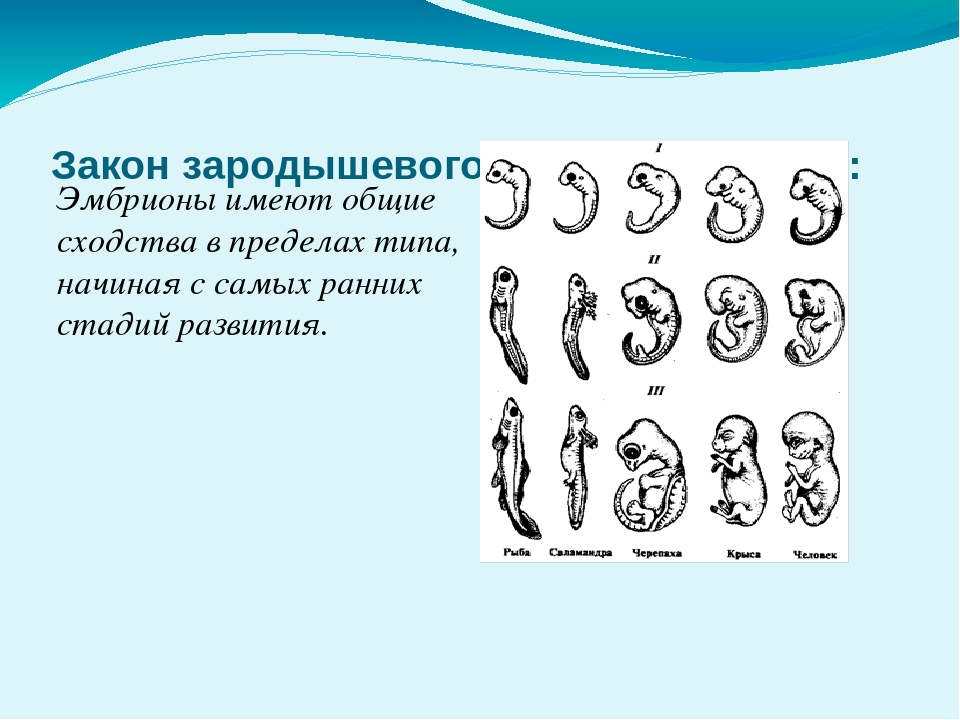 2H-J). Сходные пики индексов предков позвоночных, четвероногих и амниот указывают на то, что стадия E8.0–8.5 была наиболее консервативной на протяжении всей эволюции позвоночных. Кроме того, пик индекса предков амниот на сгруппированной стадии 14 (рис. 2J) указывает на то, что на этих стадиях экспрессируется наименьшее соотношение новейших генов развития.В свете недавнего сообщения о том, что новые гены имеют тенденцию эволюционировать быстрее, чем более старые гены [20], период E8.0-8.5 является наиболее генетически немодифицируемым или содержит самые сильные ограничения развития в эмбриогенезе мышей. Таким образом, наши анализы с генами развития показывают, что эмбриогенез мыши действительно имеет высококонсервативную стадию, характерную для предковых позвоночных, предположительно от E8.0 до E8.5 (период начала формирования глоточной дуги и сомитогенеза), что согласуется с представления о филотипической стадии позвоночных [6].Примечательно, что многие мыши с мутациями в генах развития позвоночных , экспрессированных на предполагаемой филотипической стадии позвоночных, имеют фатальные или системные фенотипы: эмбриональные или перинатальные летальные фенотипы (52%) и аномальные фенотипы нервной (33%).
2H-J). Сходные пики индексов предков позвоночных, четвероногих и амниот указывают на то, что стадия E8.0–8.5 была наиболее консервативной на протяжении всей эволюции позвоночных. Кроме того, пик индекса предков амниот на сгруппированной стадии 14 (рис. 2J) указывает на то, что на этих стадиях экспрессируется наименьшее соотношение новейших генов развития.В свете недавнего сообщения о том, что новые гены имеют тенденцию эволюционировать быстрее, чем более старые гены [20], период E8.0-8.5 является наиболее генетически немодифицируемым или содержит самые сильные ограничения развития в эмбриогенезе мышей. Таким образом, наши анализы с генами развития показывают, что эмбриогенез мыши действительно имеет высококонсервативную стадию, характерную для предковых позвоночных, предположительно от E8.0 до E8.5 (период начала формирования глоточной дуги и сомитогенеза), что согласуется с представления о филотипической стадии позвоночных [6].Примечательно, что многие мыши с мутациями в генах развития позвоночных , экспрессированных на предполагаемой филотипической стадии позвоночных, имеют фатальные или системные фенотипы: эмбриональные или перинатальные летальные фенотипы (52%) и аномальные фенотипы нервной (33%). черепно-лицевая (14%), сердечно-сосудистая (24%), дыхательная (12%) или скелетная (18%) система (см. Дополнительный файл 2: Мутантные фенотипы генов развития позвоночных, выраженные на филотипической стадии). Таким образом, многие из этих генных продуктов могут быть критическими факторами для определения фундаментальных морфологических признаков эмбрионов.Эта информация, в свою очередь, подразумевает, что мутации генов развития, экспрессируемых на предполагаемой филотипической стадии, нелегко приспосабливаются, что также подтверждает идею о том, что эта стадия имеет тенденцию быть эволюционно консервативной.
черепно-лицевая (14%), сердечно-сосудистая (24%), дыхательная (12%) или скелетная (18%) система (см. Дополнительный файл 2: Мутантные фенотипы генов развития позвоночных, выраженные на филотипической стадии). Таким образом, многие из этих генных продуктов могут быть критическими факторами для определения фундаментальных морфологических признаков эмбрионов.Эта информация, в свою очередь, подразумевает, что мутации генов развития, экспрессируемых на предполагаемой филотипической стадии, нелегко приспосабливаются, что также подтверждает идею о том, что эта стадия имеет тенденцию быть эволюционно консервативной.
Наличие двусторонней стадии
Индексы предков билатеральных и хордовых, рассчитанные с помощью генов развития, снова показали закономерности, значительно отличающиеся от таковых для позвоночных генов (рис. 2F,G против 2H-J; P <0.001), с их максимальными значениями на сгруппированных стадиях 3 и 10. Подобно результатам, полученным при анализе на уровне генома, оба этих пика были более ранними, чем предполагаемая филотипическая стадия позвоночных, охватывая период от периода дробления до начала гаструляции ( Таблица 1). Эти результаты предполагают, что этот ранний эмбриональный период несет в себе основной план тела билатеральных (то есть билатерально симметричный), что делает его «билатериотипной» стадией. Информация об экспрессии генов в эмбриогенезе Ciona кишечная также позволила нам подтвердить существование предполагаемой стадии, связанной с билатериями, в этом организме на ранней стадии эмбриогенеза (см. Дополнительный файл 3: Индексы предков, полученные для генов, экспрессированных во время эмбриогенеза Ciona кишечная ) .Наибольшее значение билатеральных индексов предков генов, экспрессируемых в ходе эмбриогенеза C. intestalis , приходится на стадию дробления и значительно превышает индексы предков более поздней эмбриональной стадии (объединенные данные экспрессии личиночной и ювенильной стадий). Однако мы не смогли получить доказательства на уровне «генов развития» из-за нехватки генных онтологий, связанных с генами C. intestalis .
Эти результаты предполагают, что этот ранний эмбриональный период несет в себе основной план тела билатеральных (то есть билатерально симметричный), что делает его «билатериотипной» стадией. Информация об экспрессии генов в эмбриогенезе Ciona кишечная также позволила нам подтвердить существование предполагаемой стадии, связанной с билатериями, в этом организме на ранней стадии эмбриогенеза (см. Дополнительный файл 3: Индексы предков, полученные для генов, экспрессированных во время эмбриогенеза Ciona кишечная ) .Наибольшее значение билатеральных индексов предков генов, экспрессируемых в ходе эмбриогенеза C. intestalis , приходится на стадию дробления и значительно превышает индексы предков более поздней эмбриональной стадии (объединенные данные экспрессии личиночной и ювенильной стадий). Однако мы не смогли получить доказательства на уровне «генов развития» из-за нехватки генных онтологий, связанных с генами C. intestalis .
Законы Бэра — Карл Эрнст фон Бэр
Законы Бэрса об эмбриональном развитии
Бэр (1828) сформулировал четыре правила.

Формулировка Рассела (1916):
(I) Общие признаки большой группы, к которой принадлежит эмбрион
появляются в разработке раньше, чем специальные символы.
(II) Менее общие структурные отношения формируются после более
общие и так далее, пока не появятся самые конкретные.
(III) Зародыш любой данной формы вместо прохождения через состояние
[т. е. конкретная конфигурация] других определенных форм [т. е. других
вида], напротив, [постепенно] отделяется от них.
(IV) По существу зародыш высшей формы животного никогда не похож на
взрослый другой формы животного [что он может делать поверхностно, например, когда
эмбрионы млекопитающих проходят стадию, когда у них появляются жаберные щели], но только
его зародыш.
Формулировка Raff & Kaufman (1983: 9):
(I) Более общие признаки большой группы животных появляются раньше
в их эмбрионах, чем более специальные символы.
(II) Из наиболее общих форм развиваются менее общие и т. д.
пока, наконец, не возникнет самое особенное.
(III) Каждый зародыш данной формы животного вместо прохождения через
другие формы, отделяется от них.
(IV) Таким образом, по существу зародыш высшей формы никогда не похож на
любая другая форма, а только ее зародыш.
Формулировка Gould (1977: 486):
Развитие идет от общего к частному. Самый ранний эмбриональный
стадии родственных организмов идентичны; добавляются отличительные черты
позже, когда неоднородность дифференцируется от однородности.Перепросмотр
невозможно; молодые зародыши представляют собой недифференцированные общие формы, не предшествующие
взрослые предки.
Формулировка Salthe (1993: 56-57):
Их можно резюмировать как закон Бэра в том смысле, что организмы
развиваются в онтогенезе, они начинают более сходными (минимально постольку, поскольку раньше
стадии более расплывчаты и поэтому неизбежно плохо различимы) и постепенно
кажутся все более и более различными по мере того, как они дифференцируются и
индивидуализировать. Иными словами, развитие идет от общего к
конкретный; происходит необратимое превращение в более высокодифференцированное
состояний, требующих все большей детализации от наблюдателя.
